

나노물질

산업 제조
현재 종양학의 과제 중 하나는 암 줄기 세포를 식별하고 특정 억제가 가능한 치료 수단을 찾는 것입니다. 이 논문은 Ehrlich 암종 세포의 표현형 특성에 대한 데이터를 종양 성장의 편리하고 따르기 쉬운 모델로 제시합니다. 에를리히 암종의 일부로서의 암 줄기세포의 증거와 CD44 + 의 중요성 및 CD44 – 이러한 유형의 종양의 성장을 유지하는 데 있어 하위 집단이 입증되었습니다. Ehrlich 암종 CD44 + 의 높은(10배) 종양 유발 활성 CD44와 비교할 경우 세포 – 세포가 입증되었습니다. 이 비교에서 CD44 + 세포는 CD44 high 의 복강 내 생성 가능성이 더 높았습니다. , CD44 + CD24 – , CD44 + CD24 + CD44 + 풀에서 암 줄기 세포의 존재를 강조하는 세포 하위 집단 세포.
이 연구에서 희토류 orthovanadates GdYVO4의 나노입자를 포함하는 합성된 하이브리드 나노복합체의 능력 :Eu 3+ 및 종양 성장을 억제하고 종양이 있는 동물의 생존을 증가시키는 콜레스테롤이 확립되었다. 종양 억제 효과에 대한 특별한 기여는 각 구성 요소에 의해 이루어집니다. Ehrlich 암종 세포를 2성분 하이브리드 복합체로 처리하면 가장 종양을 유발하는 CD44 high 농도가 최대로 감소했습니다. CD117 + 수가 동시에 증가하는 세포 대조군과 비교하여 종양 성장의 강도를 74.70 ± 4.38% 감소시킨 세포.
<섹션 데이터-제목="배경">악성 성장의 문제는 의학에서 가장 시급한 문제 중 하나로 남아 있습니다. 최근 수십 년 동안 암에 대한 새로운 치료법을 개발하는 데 약간의 진전이 있었습니다. 이것은 암에 대한 고전적 개념의 수정과 무제한 자가 재생이 가능하고 다수의 표현형 마커로 식별될 수 있는 암 줄기 세포(CSC)의 발견 때문입니다. 이들 세포의 대부분은 방사선 및 화학 요법에 내성이 있어 악성 성장 및 전이의 재발을 유발합니다. 정상 조직에 대한 손상을 최소화하면서 종양 세포를 선택적으로 비활성화할 수 있는 새로운 항암 요법 방법이 있습니다[1].
CSC는 1997년 M. Dick 팀[2]에 의해 처음으로 확인 및 설명되었습니다. 저자는 급성 골수성 백혈병을 조사했으며, 여기서 전체 세포 집단의 0.01-1%인 하위 집단이 면역결핍 NOD/SCID(비비만 당뇨병-중증 복합 면역결핍증) 마우스에 이식될 때 백혈병을 유발할 수 있습니다. 이러한 종양 유도 세포는 표현형으로 CD34 + 로 특성화되었습니다. CD38 – . 2003년 M. Al-Hajj와 M.S. Wicha는 고체 형태의 인간 유방암(BC)에서 CSC를 식별하는 데 성공했습니다[3]. 분리되지 않은 원발성 유방암 집단은 5 × 10 4 농도의 NOD/SCID 마우스에 투여했을 때 100%의 경우(10/10)에서 종양 발생 가능성을 나타내는 것으로 밝혀졌습니다. 세포/마우스. 투여되는 세포의 농도를 1 × 10 4 으로 감소 세포/마우스는 종양 형성 활성을 4배 감소시켰습니다(3/12) [3]. CD24 + CD44 + 다양한 용량으로 투여 시 분획(2 × 10 4 100 세포/마우스까지)은 종양 성장을 허용하지 않았습니다. 여기에 CD44 + CD24 – /낮은 하위 집단은 10 3 을 투여한 경우의 100%에서 종양 형성을 보여줌으로써 훨씬 더 높은 종양 형성 활성을 보였습니다. 세포/마우스. 종양을 형성하는 가장 두드러진 능력은 CD44 + 하위집단에 내재되어 있었습니다. CD24 – / 로 ESA + 표현형. 이 세포 중 단 200개를 마우스에 투여한 결과 주사 후 5개월 후에 100%(4/4)에 고형 종양이 형성되었습니다[3]. 이러한 연구는 Ponti D. et al.에 의해 계속되었는데, 이들은 유방암 생검 샘플의 특정 집단이 시험관 내에서 무혈청 배양에서 맘모스피어를 형성하는 능력을 보여주었습니다[4]. 얻어진 맘모스피어의 대부분의 세포는 CD44 + /CD24 –/낮음 SCID(중증 복합 면역 결핍증) 마우스에 투여했을 때 생체 내에서 증가된 종양 형성 가능성 뿐만 아니라 표현형. 이 하위 집단에서 종양을 형성하는 능력은 전통적으로 이식된 유방암 MCF7 라인에 비해 1000배 더 높았습니다[4]. 그러나 저자들은 CD44 + 의 20%만이 CD24 –/낮음 세포에는 자가 재생 능력이 있었습니다. 이것은 이 하위 집단의 이질성, 즉 추가 마커(ESA, ALDH)의 존재로 인한 것일 수 있으며, 이는 세포의 기능을 결정하고 또한 CD44 발현 속도와 관련될 수 있습니다. 지난 몇 년 동안 발표된 논문에 따르면 CSC는 마커의 높은 발현(CD44 high ) 가장 높은 종양 유발 활성을 갖는다[5, 6]. 5 × 10 5 의 정위 이식에서 CD44 높음 -RAS 변환 및 CD44 낮음 세포를 NOD/SCID 마우스로 변환한 결과, 하위집단 CD44 낮음 이 발견되었습니다. 낮은 종양형성(30%의 경우에서 종양이 형성됨)을 소유한 반면, CD44 높은 세포는 사례의 100%에서 종양을 형성할 수 있었습니다[6].
발표된 데이터를 요약하면 유방암 세포의 하위 집단의 분화 행은 다음과 같이 나타낼 수 있습니다.
$$ \mathrm{C}\mathrm{D}{44}^{\mathrm{high}}\to \mathrm{C}\mathrm{D}{44}^{+}\mathrm{C}\mathrm{ D}{24}^{\hbox{--}}\to \kern0.5em \mathrm{C}\mathrm{D}{44}^{+}\mathrm{C}\mathrm{D}{24} ^{+}\to \mathrm{C}\mathrm{D}{44}^{\hbox{--}}\mathrm{C}\mathrm{D}{24}^{+} $$다른 마커, 특히 Sca-1 + 이 있는 여러 세포 CSCs 단계를 주장합니다. Sca-1 녹아웃 마우스의 종양 성장 감소에 대한 데이터는 Sca-1 + 의 종양 개시 역할에 대한 가설을 지지함을 나타냅니다. 종양 형성의 초기 단계에 있는 세포[7]. 최근에 연구자들의 훨씬 더 많은 관심은 CSCs뿐만 아니라 CSCs의 보조 조절 미세 환경을 만드는 세포들에 의해 끌렸습니다. CD117 + 전통적으로 혈액 줄기 세포 풀에서 검출되는 세포는 그 중에서도 특히 주의를 기울여야 합니다[8]. 인간 유방암 세포의 총 집단은 CD117 + 이 있는 기질의 소위 암종 관련 섬유아세포로 구성됩니다. 표현형. 그들은 종양 성장을 지원하여 혈관 신생을 촉진합니다[9, 10]. Ehrlich 암종(EC) 줄기 세포 집단에 존재한다는 가정, CD44 + 의 종양 발생 가능성 연구 CD117 + 의 분수와 역할 종양 발달을 유지하는 세포에는 추가 증거가 필요합니다.
생체 내에서 CSC를 계대하기 위한 대부분의 실험은 SCID 또는 NOD/SCID 마우스에서 수행되었습니다. 이 마우스는 인간 세포의 이종이식에 대한 면역 반응에 반응하지 않습니다. 다양한 치료제의 항종양 활성을 연구하고 평가하기 위한 적절하고 적절한 실험 모델에 대한 탐색이 진행 중입니다. 그 중 하나는 생쥐의 자연발생 유방암에서 얻은 EC의 생체 내 이식 종양 세포주입니다[11]. 그러나 EC 세포의 소집단 구성 및 표현형 특성, CSC의 존재 및 이러한 유형의 종양의 성장을 유지하는 데 있어서의 중요성에 대한 간행물은 거의 없습니다. EC와 BC의 조직유전학적 유사성을 고려하면, 모의 종양의 시작 및 발달에서 암세포의 증식을 제어하는 동일한 유전자와 종양 마커 단백질의 발현을 유도하는 유사한 생화학적 경로가 관련될 수 있다고 가정할 수 있습니다. 그러나 EC 집단에서 CSC의 존재에 대한 가정과 종양 발생 가능성에 대한 연구는 추가 증거가 필요하며, 이는 본 연구의 목적 중 하나였습니다.
현재 종양학의 덜 시급한 문제는 CSC를 특이적으로 인식할 뿐만 아니라 비활성화하는 약물을 찾는 것입니다. 문제에 대한 이해라는 개념 자체가 "theranostics"(치료 + 진단) [12]라는 순간의 방향으로 형성된 기초였습니다. 진단학의 틀 내에서 암의 동시 진단 및 치료를 위한 의약품 및 도구를 사용하는 기술 접근 방식이 개발되었습니다. 진단학의 방향 중 하나는 금 나노 입자를 종양 부위에 표적화하고 광열 요법을 따르는 것입니다[13]. 종양 세포를 식별하는 다른 접근 방식은 생체 내에서 종양을 모니터링할 수 있는 강력한 광학 조영제인 양자점을 사용하는 것입니다[14]. 생체 내에서 인간 배아 줄기 세포를 비침습적으로 시각화할 수 있는 양자점의 능력은 생물 의학 적용 가능성을 입증합니다[15].
최근 희토류 원소, 즉 희토류 금속(특히 바나듐 및 그 화합물)의 나노입자(NP)로 활성화된 유전체 물질 및 광역반도체를 기반으로 하는 나노발광체에 더 큰 관심을 기울이고 있다[16]. 이 재료는 높은 광안정성, 큰 Stokes 발광의 이동, 섬광 효과의 부재 및 특징적인 좁은 발광 밴드의 안정성을 가지고 있습니다. 이와 함께 바나듐 화합물의 항종양 효과가 알려져 있습니다. 따라서, 바나듐 디클로라이드는 유사분열 이상을 유도하여 유사분열의 일시적인 억제를 유도하여 핵 이질염색질에 축적된 결과로 세포 증식을 유의하게 억제하여 후기 S에 세포 축적을 유도할 수 있는 것으로 나타났습니다. 그리고 G 2 단계 [17]. 악성 종양의 치료에 대한 유망한 것은 orthovanadates GdYVO4의 희토류 기반 NP를 기반으로 하는 하이브리드 나노복합체의 사용일 수 있습니다. :Eu 3+ 및 콜레스테롤, 우크라이나 국립 과학 아카데미의 신틸레이션 재료 연구소에서 개발 [18].
이들의 생성 목적은 표적 세포막에 친화성을 갖는 나노복합체의 조성물의 존재로 인한 항암제의 치료 효과를 향상시키는 것이었다. 하나는 생체막을 만들기 위해 암세포를 증식시켜 혈류에서 적극적으로 "철수"하는 콜레스테롤입니다. 이것은 유리 혈류 콜레스테롤과 결합할 수 있는 다수의 종양 세포 SR-B1(스캐빈저 수용체, 클래스 B 유형 I) 및 카베올린-1(Cav-1) 수용체의 표면에 존재함으로써 촉진됩니다[19] .
따라서 이 연구의 목적은 CSC의 징후가 있는 세포를 포함한 EC 세포의 하위집단 구성과 하이브리드 나노복합체로 전처리한 후의 종양 형성 활성을 확인하는 것이었습니다.
실험은 8개월 된 암컷 Balb/C 마우스에서 수행되었습니다. 마우스를 사육장의 표준 조건(실온 20 ± 2 °C, 상대 습도 50-70%, 명암 주기 12:12 h)에서 유지했습니다. 모든 실험 프로토콜은 우크라이나 하르키우에 있는 국립 과학 아카데미의 냉동생물학 및 냉동 의학 문제 연구소의 동물 윤리 위원회(2017년 1월 23일의 기록 번호 1)의 승인을 받았으며 이에 대한 유럽 협약을 준수했습니다. 실험 동물의 사용(Strasbourg, 1986), 생명 윤리에 관한 우크라이나 제1차 전국 회의(Kiev 2004)에 의해 승인되었습니다.
에를리히 암종(EC) 세포는 Balb/C 마우스의 복강(PC)에서 계대되었습니다. 복수액에 냉동보존된 EC 세포를 1차 배양으로 사용하였다[20]. 해동 후 EC 세포는 동결 해동 요인의 영향을 완화하고 자연 세포의 형태 및 기능적 특징을 얻기 위해 생체 내에서 세 번 재이식되었습니다[21]. 이에 따라 "안정화된" EC 세포를 3 × 10 6 용량으로 복강내 주사했습니다. 0.3ml의 식염수에 세포/마우스를 넣고 생체 내에서 7일 동안 배양합니다. 7일 후, 가벼운 에테르 마취하에 실험 동물을 실험에서 제거했습니다. PC의 복수액을 내경 2.69mm의 바늘을 통해 주사기로 채취하여 10ml 측정 튜브에 넣었습니다. 절대 세포 수는 Goryaev 챔버에서 계산된 EC 세포 수로 복강에 축적된 복수액(ml)의 부피를 확대하여 결정되었습니다. 총 수 최대 35.00 × 10 7 증가 7일째까지 마우스의 PC에서 EC 세포는 암 발생의 기준이었습니다[21]. 미래에는 바로 세포가 연구 대상이 되었습니다.
CD44(FITC)(no. 553133, clone IM7), CD117(FITC)(no. 553354, 클론 2B8) 및 Sca-1(FITC) no. 553333, 클론 E13-161.7) 및 CD24(PE) no. 553262, M1/69 복제) 제조업체의 지침에 따라 대조군으로서, 동일한 이소타입의 비면역 FITC- 및 PE-표지된 단일클론 항체를 첨가한 샘플("BD Biosciences"), no. 553988, 클론 A95-1 및 no. 553989, 클론 A95-1), 테스트된 마커에 대한 항체를 사용했습니다. CD44(FITC) 및 CD24(PE) 단일클론 항체를 사용하여 면역표현형 이중 염색을 수행했습니다. CD44-마커의 평균 형광이 높은 세포 10 3 (로그 척도에 따라) CD44 높음 하위 집단. "WinMDi 2.9" 소프트웨어(Joseph Trotter, La Jolla, USA)를 사용하여 결과를 기록하고 분석했습니다.
CD44 마커의 발현 수준이 높은 CSC(CD44 high ), CD44 + 의 이종 집단으로 구성됨 세포가 가장 높은 종양 형성 활성을 가지므로 자기 분류기(BDTM Imagnet)를 사용하여 전체 EC 집단에서 분리했습니다. CD44 + 를 분리하려면 제조사의 프로토콜에 따라 마커 CD44(BD, 558739) 및 이차 Mouse IgG1 Magnetic Particles-DM(BD, 557983)에 대한 1차 비표지 모노클로날 항체가 사용되었습니다. CD44 + 의 분리 순도 전체 EC 집단의 세포는 90%였습니다.
전체 인구 및 분리된 CD44의 종양 형성 능력 + 및 CD44 – EC 분획은 생체 내에서 전술한 배양 방법에 의해 비교 분석되었습니다. 실험 설정은 그림 1에 나와 있습니다.
<그림>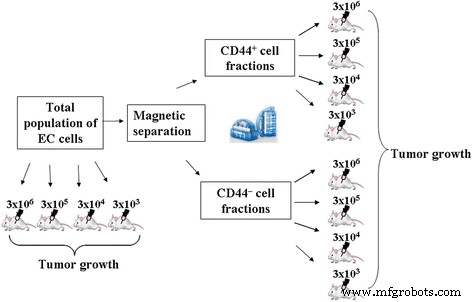
전체 모집단과 분리된 CD44 + 의 종양 발생 능력을 비교 분석한 실험 설계 및 CD44 – EC 분수
첫 번째 실험 세트에서 우리는 전체 인구의 종양 형성 능력을 평가하고 CD44 + 를 분리했습니다. 및 CD44 – EC 개시에 사용되는 표준 용량으로 동물에게 투여할 때 EC 분획(3 × 10 6 0.3ml 염수에 세포).
동물을 다음 그룹으로 나누었습니다(n =10):
그룹 1.1 - EC 세포의 총 인구 관리(3 × 10 6 세포/동물)
<리>그룹 2.1 – CD44 + 관리 EC 세포의 분획(3 × 10 6 세포/동물)
<리>그룹 3.1–CD44 관리 – 분획 EC 세포(3 × 10 6 세포/동물)
각 실험군에 접종 후 7일에 동물 PC의 총 세포 수를 세고 세포의 표현형 특성을 평가했으며(위에서 설명한 대로) CD44 높음 /CD117 + CD44 높음 비율로 결정된 EC 세포 비율 CD117에 대한 백분율 + 세포 [22]. 전체 EC 집단 및 분리된 CD44의 세포 증식 가능성 + 및 CD44 – 분획은 다음과 같은 데이터를 기반으로 추정되었습니다:배양 시간 동안 세포 집단 잉여의 다중도 인자(MF), M =N/N0; 및 시간 배가(TD), TD =(log2 2)* t /[로그2 (N /아니 0 )], 여기서 t 세포 배양 시간(h), N t에 있는 셀의 수입니다. 시각; 아니 0 초기 셀 번호[23]입니다.
두 번째 실험 세트에서는 총 모집단의 투여된 세포와 분리된 CD44 + 의 예상 최소 용량이 있었습니다. 및 CD44 – 종양 성장을 유도하는 EC 분획. 총 세포 현탁액 및 분리된 CD44 + 및 CD44 – EC 분획은 3 × 10 6 의 용량으로 마우스에 복강 내 투여되었습니다. , 3 × 10 5 , 3 × 10 4 및 3 × 10 3 0.3ml의 생리식염수에 마우스당 세포를 넣고 PC에서 7일 동안 배양합니다.
이 실험 세트에 사용된 동물은 다음 그룹으로 나뉩니다(n =10):
그룹 1.1 - EC 세포의 총 인구 관리(3 × 10 6 세포동물)
<리>그룹 1.2 - EC 세포의 총 개체군 관리(3 × 10 5 세포/동물)
<리>그룹 1.3 - EC 세포의 총 개체군 관리(3 × 10 4 세포/동물)
<리>그룹 1.4 - EC 세포의 총 개체군 관리(3 × 10 3 세포/동물)
<리>그룹 2.1 – CD44 + 관리 EC 세포의 분획(3 × 10 6 세포/동물)
<리>그룹 2.2 – CD44 + 관리 EC 세포의 분획(3 × 10 5 세포/동물)
<리>그룹 2.3 – CD44 + 관리 EC 세포의 분획(3 × 10 4 세포/동물)
<리>그룹 2.4 – CD44 + 관리 EC 세포의 분획(3 × 10 3 세포/동물)
<리>그룹 3.1–CD44 관리 – 분획 EC 세포(3 × 10 6 세포/동물)
<리>그룹 3.2 – CD44 관리 – 분획 EC 세포(3 × 10 5 세포/동물)
<리>그룹 3.3 – CD44 관리 – 분수 EC 세포(3 × 10 4 세포/동물)
<리>그룹 3.4–CD44 관리 – 분수 EC 세포(3 × 10 3 세포/동물)
모든 실험군에서 EC 접종 후 7일 후에 PC의 총 세포 수와 복수 발생 동물의 총 세포 수가 결정되었습니다.
1.30g/l 농도의 구형 나노입자(NP)(직경 2~3nm)와 농도 0.55g/l의 양 콜레스테롤("Acros Organics", 벨기에)을 포함하는 하이브리드 나노복합체는 신틸레이션 재료 연구소에서 합성되었습니다. 보고된 바와 같이 우크라이나 국립 과학 아카데미(Kharkiv)의 [18]. 희토류 원소의 orthovanadates를 기반으로 한 NP GdYVO4 :Eu 3+ 1.30g/l 농도의 구형 형태를 기술된 대로 준비했습니다[24]. 오르토바나데이트를 기반으로 하는 콜로이드 수용액은 "Cellu Sep H1" 3.5KDa 멤브레인을 사용한 투석을 통해 불순물을 정제했습니다.
하이브리드 나노복합체에서 음으로 하전된 NP는 반 데르 발스와 소수성 상호 작용으로 인해 콜레스테롤 입자의 주변을 따라 국한됩니다. NP는 정전기 상호작용을 통해 나노복합체를 안정화합니다. 합성된 나노복합체의 크기는 100nm를 초과하지 않습니다. 또한 NP는 항산화 특성을 나타내며 산화되지 않습니다. 이러한 사실은 활성산소종에 대한 콜레스테롤 수분산액의 저항성 증가에 기여한다. 하이브리드 나노복합체의 도식적 구조는 그림 2와 같다.
<사진>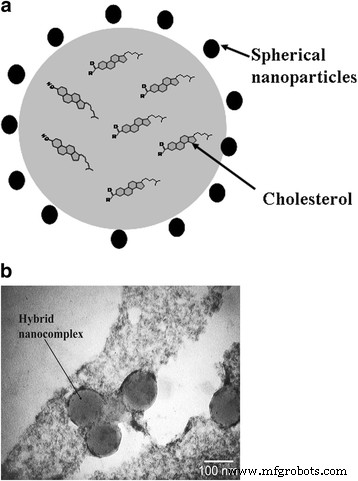
하이브리드 나노복합체:a 도식 표현 및 b 탄소 네트워크에 배치된 콜레스테롤 수용액에서 얻은 하이브리드 나노복합체의 투과 전자 현미경 사진
시험관 내 연구 동안 세포에서 하이브리드 나노복합체의 축적을 기록하기 위해 소수성 형광 염료 1,1'-디옥타데실-3,3,3',3'-테트라메틸인도카르보시아닌 과염소산염(DiI)을 콜레스테롤 수분산액에 추가로 도입할 수 있습니다. 국소 발광 분광법에서 단량체 - "J-응집체"발광 밴드 [25]의 비율에 의해 복합체가 세포막으로 통합되는 역학을 평가할 수 있습니다. 우리의 초기 연구에서는 하이브리드 나노복합체가 전체 EC 집단의 10% 이하로 통합될 수 있으며 사실상 분리된 CD44 + 의 모든 세포에 통합될 수 있음을 보여주었습니다. 발암 가능성이 가장 높은 분획. 이것은 암세포에서 나노복합체의 국부적 축적을 확인하는 방법으로 이 변형(NPs + 콜레스테롤 + DiI)에서 하이브리드 나노복합체의 사용을 허용합니다[26, 27].
하이브리드 나노복합체 또는 NP가 있는 EC 세포의 전체 현탁액을 실온에서 3시간 동안 5% 포도당 용액("Infusion" CJSC, Kyiv)에서 배양했습니다. 이러한 배양 시간은 이전에 나노복합체를 세포에 결합하는 데 최적인 것으로 밝혀졌습니다[26].
나노물질로 전처리된 EC 세포의 다음 변형이 테스트되었습니다.
옵션 1–900μl의 EC 세포(1 × 10 7 ) 100μl의 구형 NP(1.3g/l)가 추가되었습니다.
<리>옵션 2–900μl의 EC 세포(1 × 10 7 ) 100μl의 하이브리드 복합체(구형 NP(1.3g/l) + 콜레스테롤(0.55g/l))가 추가되었습니다.
대조군은 나노복합체로 처리하지 않은 5% 포도당 용액에서 배양된 EC 전체 집단의 세포였다. 각 실험군의 동물 수는 20마리 이상이었다.
배양 후, 모든 테스트 그룹의 EC 세포를 원심분리(300g에서 10분)에 의해 식염수(1:1)로 3회 세척했습니다.
나노물질 전처리 후 EC 발달의 강도는 3 × 10 6 용량의 복강내 주사에 의해 평가되었습니다. 0.3ml의 염수에 있는 세포. 모든 연구 그룹에서 EC 세포 접종 후 7일 후에 다음이 결정되었습니다.
복강 내 EC 세포의 총 수(TN).
<리>공식 Ri =(TN(c) – TN(e))에 따른 EC 성장의 억제율(Ri):TN(c) × 100%, 여기서 TN(c)–– PC의 EAC 세포 총 수 대조군, TN(e) - 실험군의 PC에 있는 EAC 세포의 총 수.
<리>EC의 성장률(Rg)은 공식 Rg(e) =Rg(c) – Ri를 사용하여 계산되었으며, 여기서 Rg(e)는 동물 실험군의 종양 성장률; Rg(c) - 대조군의 종양 성장률, Ri - 동물 실험군의 EC 성장 억제율; 대조군의 EC 성장 억제율은 100%로 하였으며, 그 때 EC 성장의 억제는 없었다.
<리>CD44 높음 /CD117 + 비율(CD44 높음 의 비율 CD117에 대한 백분율 + 셀).
<리>동물 생존은 처리되지 않은 모든 유형의 나노복합체 EC 세포로 처리된 복강내 주사 후 20일까지 평가되었습니다.
비모수 Mann-Whitney U를 사용하여 통계 처리를 수행했습니다. Statistica 6.0 소프트웨어에서 테스트합니다. 차이는 P에서 통계적으로 유의한 것으로 간주되었습니다. <0.05.
<섹션 데이터-제목="결과">얻어진 결과는 표면에 CD44, CD24, Sca-1 마커를 포함하는 EC 세포의 이질적 집단의 존재를 나타내며, 이는 미세환경(CD117)의 보조 조절 요소에 기인할 수 있습니다. 전체 EC 풀(그룹 1.1)에서 이러한 특성을 가진 세포의 농도는 표 1에 나와 있으며 EC 하위집단 구성에 대한 이전 결과와 완전히 일치합니다[28]. 거의 모든 EC 세포에서 Sca-1 구조의 식별은 이를 이 종양 유형의 다용도 마커로 간주할 수 있습니다.
CSC의 표현형 식별 측면에서 가장 유익한 것은 CD44 분자 발현이며, 이는 자체적으로 또는 다른 표면 마커와 함께 EC를 비롯한 다양한 종양에서 이 세포 집단을 분리하는 데 사용됩니다. 고전적인 개념에 따르면 유방암 발병 중 종양 세포의 분화는 CD-44 수용체의 발현 감소와 함께 CD24 마커를 발현하는 세포의 점진적인 소멸 및 출현을 동반합니다[3].
EC 동안 CSC의 역할 후보는 CD44 높은 세포일 수 있습니다. CD44 + 의 일부인 표현형 CD24 – - 인구. CD44 + 하위집단의 기능적 활동에 대한 EC의 의존성에 대한 이 가정 세포는 CD44 + 에 의해 유도된 종양 성장의 강도를 평가할 때 테스트되었습니다. 및 CD44 – 파벌 및 EC 총 인구. 표 1은 가장 높은 종양 유도 활성이 CD44 + 의 세포에 내재되어 있음을 보여줍니다 분수. 실제로 3x10 6 관리 후 CD44 + 세포(그룹 2.1)에서 PC의 절대 세포 수는 전체 EC 세포(그룹 1.1)보다 23배 더 많았고 CD44 – 보다 105배 더 많았습니다. 분획을 투여하였다(그룹 3.1).
이로써, 발달하는 종양의 양적 구성뿐만 아니라 질적 구성의 변화도 발견되었다. CD44 + 의 비율 CD44 + 의 주요 함량으로 복수를 형성했습니다. 세포, 즉, CD44 높음 , CD44 + CD24 – 및 CD44 + CD24 + 세포. 게다가, CD44의 농도 높음 세포는 그룹 1.1에 비해 2배, 그룹 3.1에서 16배 더 높았다. CD44의 비율 – , 대조적으로, 더 성숙한 세포, 즉 CD44를 가진 세포를 포함하는 종양을 형성했습니다. – CD24 + 표현형. 그룹 3.1에서 세포의 아집단 구성의 바로 재분배가 PC에서 세포의 최소 절대 함량을 분명히 결정했습니다.
중요한 것은 CD117 + 을 가진 하위집단의 EC 세포 사이에 존재한다는 확립된 사실입니다. 채점자. CD117의 분자는 막횡단 티로신 키나제 수용체입니다. 정상적인 조건에서는 해당 리간드, 즉 줄기 세포 성장 인자(SCGF)에 의해 활성화됩니다[29]. 종양학적 병리학에서 c-KIT 수용체의 리간드-의존적 활성화가 발생하며, 가장 흔히(최대 92%의 경우) c-kit 종양유전자 돌연변이의 결과이거나 이 수용체 기능의 조절 메커니즘의 장애로 인해 발생합니다. [30].
CD117 + 고려 중 종양 미세 환경의 세포로서의 세포 CD117 + 의 존재 또는 부재에 대한 종양 성장 강도 의존성의 확립된 사실 세포 및 CD44 높음 과의 농도 관계 세포는 논리입니다. 표 1에서 알 수 있듯이 PC에 전체 세포 집단(그룹 1.1.)을 도입하여 EC를 시작할 때 34.80 ± 1.27 × 10 7 이 형성되었습니다. CD44 높은 세포 /CD117 + 비율은 0.02 상대 단위와 같습니다.
CD44의 종양 유발 가능성 – 비율은 4배 낮았으며(그룹 3.1) CD44 높음 감소로 나타났습니다. /CD117 + 그룹 1.1과 비교하면 동일한 정도(4배)의 비율. 이 변화는 CD44 높음 /CD117 + 지수는 주로 CD44 high 의 감소로 인한 것입니다. CD117 + 의 감소된 함량의 배경에 농도(8.5배) 세포(2회)도 마찬가지입니다.
CD44 + 에 의해 생성된 복수 성장의 강도를 평가할 때 분획, PC의 총 세포 수의 상당한 증가(그룹 1.1과 비교하는 경우 거의 24배)가 주목되었습니다. 또한 중요한 것은 CD44 high 의 2배 초과입니다. CD117 + 의 집중 및 결핍 세포. CD44 + 의 초기 자료에서 데이터의 유세포 분석에 따른 분획(배양 전), CD44의 함량 높음 세포는 EC 세포의 전체 집단보다 15배 더 높았다(데이터는 제시되지 않음).
위에서 언급한 내용을 요약하자면, EC의 개시에서 중요한 역할은 CD44 마커의 높은 발현율(CD44 high ), CD117 + 의 가장 중요한 기능 중 하나입니다. 하위 집단은 CD44 높은 종양 형성 활성의 조절("억제")입니다. 세포. CD117 + 부재 세포(그룹 2.1)는 전체 CD44 풀의 증식 및 분화 가능성을 증가시키는 것으로 보입니다 + PC의 총 세포 수를 크게 증가시킵니다.
전체 EC 세포 풀 및 CD44 + 의 증식 가능성 분석 파벌은 이 해석을 선호합니다. 그룹 2.1의 PC에서 배양된 전체 인구의 곱셈 인자(MF)가 나타납니다. 7일 동안 그룹 1.1에 비해 거의 24배 증가했습니다. 이는 CD44 + 에서 성장한 복수 세포 집단을 특징지을 수 있는 그룹 2.1의 1.1–14.70 ± 1.35 시간에서 2배의 세포 시간 감소를 동반했습니다. 더 활발하게 증식하는 분수(표 1).
CD44 + 의 특별한 역할을 증명하기 위해 최소 용량으로도 EC를 투여할 때 종양의 개시 및 유지에 있는 세포, 분리된 CD44 + 의 종양 형성 능력을 비교 평가하는 것이 중요했습니다. 및 CD44 – 다양한 농도로 투여할 때 분획. 3 × 10 6 의 도입 후 전체 EC 집단의 세포에서 종양 성장은 동물의 100%에서 관찰되었습니다(10/10)(표 2). 투여되는 세포의 10배 용량 감소(3 × 10 5 ) resulted in a proportional decrease in absolute number of cells in the PC, tumor developed only in 50% of animals (Table 2). Reducing the administered dose of total EC population of cells down to 3 × 10 4 did not lead to tumor formation in the PC.
Initiations of EC by introducing of CD44 + cells at concentrations of 3 × 10 6 및 3 × 10 5 cells per animal resulted in almost 100% tumor development for both cases. Herewith, tumorigenic potential of CD44 + fraction exceeded that of total population of EC cells administered in the same doses (in 23 and 21 times, respectively). Moreover, introduction of 3 × 10 4 cells of CD44 + fraction caused a tumor formation in 33% of animals, while total population of EC cells used in the same dose, did not cause the formation of ascites. With the introduction of 3 × 10 3 cells of CD44 + fraction, no animals with the developed EC have been identified.
Fraction of CD44 – cells just in a dose of 3 × 10 6 was capable of forming tumors in 50% of animals, the number of cells in the PC in this case was 4.5 times less than when introducing the total population and in 105.9 times less than when inducing by CD44 + fraction. Thus, the results of this part of research suggest that CSCs are mainly present in the pool of cells with CD44 + phenotype. This emphasizes the importance of this subpopulation of cells in initiation and development of EC.
As noted above, identification and inactivation of CSCs is a major theoretical and practical issue of oncology. On this basis, the next task of our study was to investigate the impact of hybrid nanocomplexes designed at the Institute for Scintillation Materials of National Academy of Sciences of Ukraine on the tumorigenic activity of EC cells.
As Table 3 demonstrates an incubation of EC cells with only NPs as a component of hybrid nanocomplexes (option 1) decreased the concentration of CD44 high virtually twice if compared to the control and 5 times the content CD44 + CD24 – cells in ascites formed in vivo. The number in it of more differentiated CD44 + CD24 + , CD44 – CD24 + cells remained practically unchanged if compared to the control. In this group, there was established reduction of CD117 + cells (35%) at a slightly changed content of Sca-1 + subpopulation. Based on the data, the inhibition rate of EC growth (59.41 ± 3.45%) in variant 1 was accompanied by a twofold decrease in the concentrations of CD44 high cells in comparison with the control that was also reflected in the reduction of CD44 high /CD117 + ratio (Table. 3).
Pretreatment of EC cells with hybrid nanocomplexes (option 2) reduced almost 10 times the concentration of CD44 high and CD44 + CD24 – cells in the developed ascites if compared to the control (Table 3). It should be noted that the concentration of more differentiated CD44 + CD24 + and CD44 – CD24 + cells after this treatment increased slightly if compared to the control. The redistribution pattern of EC subpopulation composition in this option was accompanied with a pronounced enhancement of tumor growth inhibition compared to option 1 (74.70 ± 4.38 and 59.41 ± 3.45%, respectively, P < 0.05) that underlined the importance of cholesterol as a targeted compound of antitumor therapy. Pretreatment with hybrid nanocomplexes (option 2) led to maximal reduction there was found a maximum reduction of CD44 high /CD117 + ratio (10 times) as compared with option 1, that again confirmed a specific role of ratio of these cell subpopulations in the EC growth.
For all the types of EC pretreatment, the reduction of CD44 high /CD117 + ratio was accompanied by a decrease in tumor growth rate and increased survival of animals to day 20 of EC development (Fig. 3).

Tumor growth rate of EC, survival of animals and CD44 high /CD117 + ratio after incubation with nanocomplexes. Note:differences are statistically significant as compared with administration of the control (*), option 1 (**) (P < 0.05)
One of the tasks of current oncology is elucidation of the mechanisms of initiation and development of malignant neoplasms. Mandatory participants in these events are the CSCs and so-called accessory-regulatory cells of tumor microenvironment. The variety of functional and structural characteristics of the CSCs in the development of different types of tumors determines the need for their further study. This is facilitated by the expansion of experimental model systems. One of them is the transplantable line of tumor cells of EC.
The elucidation of the peculiarities of this experimental model development, the subpopulation composition of tumor and tumorigenic potential of individual cell populations within the general pool of the EC cells will facilitate the development of new approaches to cancer therapy.
Using the method of phenotypic evaluation of progenitor cells of various levels of differentiation in the tumor focus makes it possible the identifying the stages, dynamics of development and invasiveness of the process. The established fact of heterogeneity of the EC subpopulation composition is important and there has been emphasized the value of CD44 + subpopulation in maintaining the growth of this type of tumor.
The most important role in implementing a tumorigenesis is played by an expression rate of the molecule. Indeed, in contrast to leukocytes for adhesion of those normally a low expression rate of CD44 receptor is required, triggering and self-maintenance in CSCs are implemented its much greater density on a cell surface [31].
It is known that CD44-glycoprotein is a hyaluronic acid (HA) receptor, a main component of extracellular matrix. The emerging set of HA-CD44 activates many receptor tyrosine kinases, resulting in activation of PI3K/Akt/ mTOR way [32, 33], which plays the role of a single universal signal transmission mechanism to the translation apparatus and is responsible for the integration of proliferative stimuli.
Among two known CD44-isoforms in normal hematopoietic cells its standard isoform (CD44s) is predominantly expressed [34]. In most malignant tissues there were detected both CD44s and variable isoforms of CD44- molecule (CD44v), resulting from alternative splicing of exons 6-15. Namely alternative splicing leads to a lengthening of CD44-extracellular domain, promoting its greater interaction with HA and tumor metastasis [35]. Due to that the role of CD44 high cells in triggering and maintaining the tumorogenesis is clear. It was previously found that a minor subpopulation of CD44 high cells had a high proliferative potential and played a critical role in EC developing [20].
In this paper, a special role of CD44 + -cells of the EC in initiation and maintenance of the tumor process in the EC under administration even in minimal doses has been shown. CD44 + cells were able to form a tumor even at a cell concentration of 100 times lower (10 4 cells/ mouse) if compared with the introduction of a total EC population (10 6 cells / mouse). The belonging of tumor cells to the CD44 + fraction was also confirmed by the fact that the EC initiation by the fraction of CD44-cells even at a dose of 10 6 cells / mouse caused the formation of a tumor only in 50% of cases, with an absolute number of cells in the PC 5 times less than in under introduction of a similar amount of the total population of EC and more than 100 times less than after the introduced CD44 + -fraction.
This is in accordance with the data of Shipitsin M et al. has shown that CD44 + and CD24 + cells in breast cancer development there are cell populations with different genetic profiles [36]. The research performed by Shipitsin M CD24 + cells have been noted to be more differentiated, while more progenitor-like functions are inherent to CD44 + cells. The research performed by Shipitsin M CD24 + cells have been noted to be more differentiated, while more progenitor-like functions are inherent to CD44 + cells. The authors suggest that CD24 + cells can be derived from CD44 + cells [36]. Fillmore C. and Kuperwasser C. supposed that CD24 + population was mainly characterized by less differentiated basal type of breast cancer, and CD44 + cells caused the development of luminal form of breast cancer, being more differentiated type of tumor [37].
Analyzing the patterns of tumor development, the classic hypothesis of «seed and soil» looks very actual [38], which postulated that an appropriate microenvironment (soil) is required for optimal growth of tumor cells (CSCs).
Most often the carcinoma-associated fibroblasts (CAFs) act as a tumor stroma in breast cancer and pancreatic cancer [39]. It has been shown that the CAFs, derived from invasive forms of human breast carcinomas, activated much stronger the growth of human breast cancer cell line MCF-7-Ras when administered to immunodeficient mice if compared with normal fibroblasts [9]. This function is implemented by the microenvironment cells due to the secretion by them of cytokines, chemokines and growth factors [10, 40].
Although so far the phenotypic identification of the microenvironment cells for various types of tumor has remained a subject of debate, most often used for this purpose the surface markers of primitive hematopoietic and endothelial cells, including c-kit (CD 117), CD133, VE-cadherin, VEGFR-2 and endoglin are used [41]. In this experimental model the most probable candidate to the role of tumor microenvironment cells is CD 117 + .
It is known that the c-KIT receptor (CD117 + ) is highly expressed in normal epithelium of the breast and progressively decreases with the development of breast carcinoma in situ and is almost completely lost in invasive breast cancer [42, 43]. Some authors proposed this kind of change in the expression rate of this marker as a possible test to assess the effectiveness of antitumor therapy [44].
Previously, after analysis of the significance of the content ratios for different subpopulations of EC cells when maintaining tumor growth, we proposed to use the CD44 high /CD117 + ratio as a prognostic criterion of tumor development [22].
Adequacy of using this index is confirmed in this study using the applied nanocomposites as therapeutic agents when treating the EC. The inhibition rate of EC growth (59.41 ± 3.45%) when treated with spherical NPs (option 1) was accompanied by a 2-fold decrease if compared to the control in the CD44 high -cell concentration, which was reflected in the reduced CD44 high / CD117 + 인덱스. The maximum decrease in the CD44 high /CD117 + index (10 times if compared to option 1) was established using the hybrid nanocomplexes for a pre-treatment of EC cells. Thus, many cells of a total pool of EC, but primarily those with the phenotype CD44 high and CD117 + , can be the target of the effect of the studied nanocomplexes (both direct and indirect). A significant decrease in their concentrations in the growing pool of EC after pretreatment with hybrid nanocomplexes clearly coincides with a reduced intensity of tumor growth.
Judging by the decrease in the amount of CD44 high as the most potent CSCs forming the entire subsequent series of advanced tumor cells, the main component in manifestation of antitumor effect of the synthesized hybrid nanocomplexes is spherical NPs. Introduction of cholesterol having affinity to tumor cell membranes into composition of hybrid nanocomplexes enhanced an inhibitory activity of NPs. Similar data were obtained by Betker J.L. et al. after analysis of the structure and functioning principles of the membranes of tumor cells. The authors concluded that the incorporation of cholesterol into membranes of tumor cells could be a prerequisite for a targeted delivery of liposomes with therapeutic agents directly into a cell.
Thus, the importance of cooperative interactions of cells with different phenotypic signs in maintaining the EC growth has been proven. The cells with the CD44 high phenotype being the part of the population of CD44 + CD24 – can be considered as CSCs in this model system. The use of new forms of nanocomposites that are capable to bind to CSCs and induce tumor destruction as the EC is a promising direction the treatment of oncopathology.
On the base of the findings of phenotypic assessment and functional potential studies, the Ehrlich carcinoma is a heterogeneous population of tumor cells of varying differentiation extent referred to high and less potent tumor-inducing precursors, as well as the cells composing their microenvironment.
A high (tenfold) tumorigenic activity of the EC CD44 + cells if compared to CD44 – cells was proven. In this pair of comparison, the CD44 + cells had a higher potential of generating in PC of CD44 high , CD44 + CD24 – , CD44 + CD24 + cell subpopulations, highlighting the presence of CSCs in a pool of CD44 + cells.
There was found an ability of the synthesized nanocomplexes based on rare earth orthovanadates and cholesterol to inhibit the growth of CD44 + cell pool (CD44 high , CD44 + CD24 – , CD44 + CD24 + ) that was accompanied by a reduced intensity of EC growth (by 75%) and increased survival of the animal with tumors (in 3.5 times) in comparison with the control.
It has been shown that the reduction in tumor growth rate after pretreatment with hybrid nanocomplexes was accompanied with a change in the composition of EC subpopulation that was reflected in a decrease in the CD44 high /CD117 + ratio. This ratio can be offered as one of diagnostic and prognostic tests of the severity and extent of oncology inactivation.
Breast cancer
Carcinoma-associated fibroblasts
Cancer stem cells
1,1′-Dioctadecyl-3,3,3′,3′-tetramethylindocarbocyanine perchlorate
Ehrlich carcinoma
Hyaluronic acid
Multiplicity factor
Nonobese diabetic-severe combined immunodeficiency mice
Nanoparticles
Peritoneal cavity
Growth rate of EC
Inhibition rate of EC growth
Severe combined immunodeficiency mice
Time doubling
나노물질
초록 온열요법은 비침습성, 부작용 및 독성 최소화, 시행 용이성으로 인해 암 질환을 치료하는 가장 환자 친화적인 방법 중 하나이며 광열 유발 용량 시스템과 같은 새로운 치료 방법의 개발을 촉진합니다. 여기에서 이 연구는 Cs0.33의 광열 효과 변수를 조사합니다. WO3 시험관 내에서 HepG2 간암 세포주에 대한 나노입자(NP), 조사 기간, 광학 전력 밀도 및 NP 농도는 근적외선(NIR) 조사 열 용량의 공식화로 이어집니다. 명시적으로, 120nm의 입자 형상 크기를 갖는 NP는 일련의 산화-환원(REDOX) 반응, 열 어닐
초록 파킨슨병(PD)은 중뇌에서 도파민성 뉴런의 점진적인 소실을 특징으로 하며, 줄기 세포 이식 방법은 치료를 위한 유망한 전략을 제공합니다. 이러한 연구에서 생체 내 이식된 세포의 생물학적 행동을 추적하는 것은 줄기 세포 기능에 대한 기본 이해와 임상 효과 평가에 필수적입니다. 본 연구에서 우리는 열분해 방법과 2단계 개질을 통해 폴리아크릴산(PAA)과 메톡시폴리에틸렌글리콜아민(PEG)으로 코팅된 새로운 초소형 초상자성 산화철 나노입자를 개발했습니다. USPIO-PAA/PEG NP는 TEM 및 FTIR에서 볼 수 있듯이 10.0