

나노물질

산업 제조
패혈증 유발 면역 억제는 치료 실패의 주요 특징 중 하나로 인식됩니다. 주로 억제 특성이 특징인 골수 유래 억제 세포(MDSC)는 패혈증에서 확장되는 것으로 보고되었습니다. FDA 승인 철분 보충제인 FMT(Ferumoxytol)는 종양에서 면역 조절 특성을 갖는 것으로 나타났습니다. 그러나 FMT가 후기 패혈증 면역억제를 감소시키기 위해 MDSC의 기능을 변경하는지 여부는 불분명합니다. 여기에서 우리는 패혈증의 후기 단계에서 지질다당류(LPS) 유도 면역 억제를 개선하기 위해 MDSC에 대한 FMT의 면역 조절 효과를 보여주었습니다. 내재화된 FMT로 세포를 분리하고 세포내 철 함량을 검출한 결과 MDSC가 FMT를 흡수할 수 있음을 보여주었습니다. 낮은 용량의 FMT는 MDSC의 세포 생존력에 영향을 미치지 않았지만 FMT는 시험관 내에서 MDSC의 확장을 억제했습니다. 또한, FMT는 MDSC에서 Arg-1, S100A8, S100A9 및 p47phox의 발현 수준과 ROS 생산을 유의하게 하향 조절했습니다. FMT는 과립구 MDSC(G-MDSC)의 비율을 감소시키고 MDSC의 대식세포로의 분화를 촉진했습니다. 또한, FMT는 간 기능 장애의 일부 생화학적 지표뿐만 아니라 폐 및 간 괴사 부위의 백혈구 모집 및 폐포벽 비후를 감소시켰습니다. FMT는 LPS로 유도된 패혈증 마우스의 비장에서 G-MDSC와 단핵구 MDSC(M-MDSC)의 비율을 감소시켰습니다. 참고로, FMT는 G-MDSC와 M-MDSC 모두의 T 세포 면역억제 기능을 감소시켰습니다. 예상대로 FMT는 또한 비장 CD11b + 에서 Arg-1 및 p47phox 유전자 발현을 유의하게 감소시켰습니다. 1학년 + LPS-챌린지 마우스에서 분리된 세포. 이러한 데이터는 FMT가 Arg-1 및 ROS 생성을 감소시켜 MDSC의 면역억제 기능을 감소시켰음을 나타내며, 이는 FMT가 패혈증 말기의 장기간 면역억제를 감소시킬 수 있음을 시사합니다.
FMT는 식품의약국에서 승인한 철분보충제로 성인 만성신장질환(CKD) 환자의 철결핍성 빈혈 치료에 사용되어 왔으며[1], FMT는 조영제 및 약물 운반체로도 널리 사용되고 있다[2]. . 이전 연구에서는 FMT가 높은 CD86 + 을 향한 M2 대식세포의 표현형 이동을 유도하는 능력과 같은 면역 조절 특성을 가지고 있음을 보여주었습니다. , TNF-α 양성 M1 대식세포 아형[3, 4]. 그러나 FMT가 다른 면역 세포에 미치는 영향은 조사되지 않았습니다.
MDSC는 강력한 면역 억제 능력을 가진 미성숙 골수 세포의 이질적인 집단입니다[5]. 마우스에서 MDSC는 Gr-1 + 으로 식별됩니다. CD11b + 인간 MDSC는 Gr-1 상동체가 없고 CD14 - 로 정의됩니다. HLA-DR − CD11b + CD33 + 또는 CD14 + HLA-DR − CD11b + CD33 + 세포 [6]. MDSC는 과립구 또는 PMN-MDSC와 M-MDSC의 두 가지 큰 세포 그룹으로 구성되며 두 집단 모두 iNOS, ROS 및 Arg-1의 생산을 통해 면역 억제 기능을 가지고 있습니다. Arg-1의 활성 증가는 l-arginine을 고갈시키고 l-arginine의 부족은 CD3 zeta-chain의 발현을 감소시켜 T 세포 증식을 억제합니다. INOS는 T 세포에서 JAK3와 STAT6의 기능과 MHC II의 발현을 억제하는 NO를 생성합니다. ROS는 T 세포 수용체의 번역 후 변형을 유도하고 항원 특이적 T 세포 무반응을 유발할 수 있습니다[7]. M-MDSC는 단핵구 전구체에서 발생하며 적절한 사이토카인 조건에서 대식세포와 DC로 분화할 수 있습니다. MDSC에 대한 대부분의 지식은 암 연구에서 비롯됩니다. 최근에는 염증, 화상, 자가면역질환 등의 급성 및 만성 염증성 질환에서도 MDSCs의 확장이 보고되고 있다[8, 9]. 최근에 쥐의 식도와 비장에서 MDSCs에 의한 높은 수준의 형광 나노입자 흡수가 영상 연구에서 보고되었습니다[10]. 그러나 FMT가 염증성 질환의 발병에 관여하는 MDSC의 기능을 변경하는지 여부는 불분명합니다.
생명을 위협하는 장기 기능 장애를 유발할 수 있는 심각한 감염에 대한 조절되지 않는 숙주 염증 반응인 패혈증은 중환자실에서 가장 흔한 사망 원인입니다. 패혈증은 초기에 치료하지 않으면 빠르게 장기적인 면역억제로 변하는 압도적인 전염증 반응을 시작합니다. 패혈증 환자[12,13,14]뿐만 아니라 패혈증 동물 모델[8, 11]의 비장과 림프절에서 MDSC의 확장이 설명되었습니다. MDSC와 같은 면역 억제 세포의 활성화는 선천성 면역 시스템의 과민 반응을 적절하게 제어하는 데 중요하지만, 과도한 확장은 과면역 억제로 이어질 수 있으며, 이는 후기 패혈증에서 2차 감염 및 사망의 주된 원동력입니다. MDSC의 증가된 수는 림프구 증식의 억제와 상관관계가 있습니다. MDSC는 골수에서 표현형을 획득한 다음 2차 림프 기관으로 이동하여 T 세포 반응을 억제하고 LPS 면역 억제 마우스에서 증식을 억제하는 것으로 보고되었습니다[8]. 장기간의 패혈증이 있는 대부분의 환자는 면역억제 상태를 나타내며 대부분의 패혈증 사망률은 후기 패혈증 면역억제에 기인합니다. MDSC를 제거하면 면역 반응을 회복하여 후기 패혈증에 유익한 효과가 있을 수 있습니다[15]. 따라서 우리는 FMT가 MDSC의 기능을 조절하여 후기 패혈증 면역 억제를 감소시킬 수 있다고 가정했습니다.
여기에서 우리는 MDSC에 대한 FMT의 면역 조절 효과를 보여주었습니다. 우리는 MDSC가 시험관 내에서 FMT를 식균할 수 있음을 밝히기 위해 FMT의 자기적 특성을 활용했습니다. 또한, 우리는 FMT가 Arg-1 및 ROS의 하향 조절을 통해 MDSC의 면역 억제 기능을 약화시킨다는 것을 발견했습니다. 또한, 생체 내 실험에서 FMT가 T 세포를 억제하는 MDSC의 능력을 약화시켜 마우스에서 LPS 유도 패혈증의 후기 단계를 개선했음을 확인했습니다. 우리의 데이터는 FMT의 면역 조절 기능이 패혈증 후기에 발생하는 면역 억제를 치료하는 데 사용될 수 있음을 시사합니다.
수컷 C57BL/6 마우스(8-10 주령)는 Nanjing University(Nanjing, China)의 Model Animal Research Center에서 구입했습니다. 마우스는 24시간 명암 주기의 특정 무병원체(SPF) 조건에서 사육되었으며 음식과 물에 자유롭게 접근할 수 있었습니다. 쥐에 대한 모든 실험은 Nanjing University Institutional Animal Care and Use Committee의 승인을 받았습니다.
세포 내 FMT 분포는 제조사의 지침에 따라 Perl의 Prussian blue 염색 키트(Solarbio, China)를 사용하여 검출되었습니다. 간단히 말해서, 세포를 4% paraformaldehyde에서 15분 동안 고정한 다음 10% ferrocyanide 칼륨과 25-30분 동안 배양한 다음 10분 동안 핵 패스트 레드로 세척하고 대조염색합니다. 세포질에 파란색 입자가 포함된 세포는 프러시안 블루 염색에 양성이었습니다.
골수 세포는 야생형 마우스의 대퇴골 및 경골을 플러싱한 후 RBC 용해로 수득하였다. 골수 세포를 10% FBS, 1% v/v 페니실린 및 스트렙토마이신(Gibco, CA), 40 ng/mL 과립구-대식세포 집락 자극 인자(GM-CSF)(Miltenyi Biotech, 독일) 및 40 ng/mL IL-6(Miltenyi Biotech, 독일), 37 °C, 5% CO2 4 일 동안 인큐베이터. 추가 실험을 위해 비부착 세포를 수집했습니다.
세포 생존율은 제조업체의 지침에 따라 Cell Counting Kit-8(Dojino, Kumamoto, Japan)을 사용하여 분석되었습니다. 간단히 말해서, MDSC(5 × 10 3 )를 96웰 배양 플레이트에 플레이팅하고 5% CO2에서 배양했습니다. 처리 전 37 °C에서 대기. 그런 다음 배지를 다양한 농도의 FMT(250, 500, 1000, 2000 μg/mL)를 포함하는 새로운 배지로 24시간 동안 교체했습니다. 이어서 상층액을 버리고 CCK-8 용액을 함유하는 새로운 배지를 첨가하였다. 37 °C에서 3시간 동안 배양한 후 Gen5 마이크로플레이트 리더(BioTek, USA)를 사용하여 450 nm에서 흡광도를 측정했습니다.
TRIzol 시약(Invitrogen, USA)을 사용하여 세포에서 총 RNA를 추출하고 Nanodrop Spectrophotometer(Thermo Scientific, USA)로 정량한 후 HiScript II 1st strand cDNA Synthesis Kit(Vazyme Biotech Co., Ltd, China)를 사용하여 역전사했습니다. 제조업체의 지침에 따라. 실시간 정량적 중합효소 연쇄 반응(PCR)은 Applied Biosystems StepOne-Plus 실시간 qPCR 시스템(Applied Biosystems, Forster City, CA)에서 SYBR Green PCR Kit(Bio-Rad, Hercules, CA)를 사용하여 수행되었습니다. 유전자 발현은 2 -ΔΔCt 를 사용하여 GAPDH로 정규화되었습니다. 방법. 프라이머 서열은 추가 파일 1:표 S1에 나열되어 있습니다.
조직은 Hanks' Balanced Salt Solution(HBSS)에서 37 °C에서 30분 동안 콜라게나아제 유형 D(1 mg/mL) 및 DNase I(0.1 mg/mL)을 포함하는 단일 세포 현탁액으로 준비한 다음 비장이 용해되었다. 세포 현탁액을 70μm 세포 여과기를 통해 여과하고 림프구를 4 °C에서 5분 동안 300 g에서 원심분리하여 수집했습니다. 세척 후, 세포는 형광 활성화 세포 분류를 위해 즉시 준비되었습니다. 단일 세포를 FcR 차단 시약(Miltenyi Biotech, Germany)과 함께 10분 동안 인큐베이션한 후 다음 항체 접합체로 20분 동안 염색했습니다:FITC-표지된 항-CD11b, 항-CD11c, 항-CD4 및 항-CD8; PE-표지된 항-Ly6G; APC-표지된 항-Ly6C, 항-CD3, 항-F4/80 및 항-MHC II(BioLegend, CA). 세척 후 FACS Calibur(BD, CA)로 세포를 검출하고 FlowJo 7.6(Tree Star, Inc., USA)을 사용하여 데이터를 분석했습니다.
간 및 폐 조직은 4% 인산염 완충 포름알데히드로 고정되었습니다. 고정된 조직을 파라핀에 포매하고 5μm 두께의 절편을 광학현미경을 위해 헤마톡실린과 에오신으로 염색하였다.
TNFα, MCP-1 및 IL-1β의 혈청 수준은 제조업체의 지침에 따라 특정 ELISA 키트(Dakewe, China)를 사용하여 결정되었습니다. 각 샘플은 중복 실행되었습니다.
세포 내 ROS는 2,7-dichlorodihydrofluorescein diacetate(DCFH-DA) 키트(Beyotime, China)를 사용하여 제조업체의 지침에 따라 측정되었습니다. 간단히 말해서, 세포를 PBS로 2회 세척하고 5μM DCFH-DA와 함께 37℃, 암실에서 30분 동안 인큐베이션한 다음, RPMI-1640 배지로 세척하고 PBS에 재현탁시켰다. 세포는 유세포 분석기(BD, CA)를 사용하여 세포 내 녹색 형광에 대해 분석되었습니다.
총 CD3 + CD3ε MicroBead 키트(Miltenyi Biotech, Germany)와 배양된 CFSE 염색(Invitrogen, USA)을 사용하여 야생형 마우스의 비장에서 T 세포를 농축했습니다. 항-CD3/CD28 항체-유도된 T 세포 증식을 위해, T 세포는 항-CD3 항체(1 ㎍/mL) 및 항-CD28 항체(1 ㎍/mL)로 활성화되었다. G-MDSC(Gr-1 높음 Ly-6G + ) 및 M-MDSC(Gr-1 dim Ly-6G − ) 제조사 지침에 따라 골수 유래 억제 세포 분리 키트(Miltenyi Biotech, Germany)를 사용하여 FMT 처리 또는 비히클 처리 마우스의 비장에서 분리했습니다. MDSC는 CFSE로 표지된 CD3 + 와 1:1 비율로 공배양되었습니다. 3 일 동안 96웰 평평한 바닥 플레이트의 T 세포. CFSE는 각 분할에 따라 딸 세포 간에 균등하게 분할됩니다. 따라서 증식은 유세포 분석에 의해 결정되었습니다. 분류 후 세포의 순도는> 90%였습니다.
모든 통계는 GraphPad Prism 6 소프트웨어(GraphPad Software, CA)를 사용하여 계산되었으며 평균 ± 표준 편차(SD)로 표시됩니다. 두 그룹 간의 차이는 Mann–Whitney U에 의해 분석되었습니다. 테스트 또는 짝을 이루지 않은 양측 학생의 t 테스트. 일원 ANOVA는 여러 그룹의 비교에 사용되었습니다. 모든 실험은 적어도 세 번 반복되었습니다. p와의 차이점 값 <0.05는 통계적으로 유의한 것으로 간주되었습니다.
<섹션 데이터-제목="결과">FMT가 내재화된 세포가 MACS MicroBeads에 의해 분리되는지 확인하기 위해 24시간 동안 FMT(1000 ng/mL)로 처리된 대식세포에서 세포내 철 함량을 검출하기 위해 프러시안 블루 염색을 사용했습니다. 세포는 자기 분리 이전에 자기적으로 선택된 FMT 양성 세포(FMT+)와 자기적으로 분리되지 않은 세포(FMT-)의 세 그룹으로 나눴습니다. 프러시안 블루 염색은 자기적으로 선택된 대부분의 세포가 프러시안 블루 양성임을 보여주었다(그림 1a). MDSC, 대식세포 및 DC 사이에서 FMT를 식균하는 능력을 비교하기 위해 FMT로 6시간, 12시간, 24시간 및 48시간 동안 처리한 순진한 C57BL/6 마우스에서 비장세포를 분리했습니다. 비장 세포는 자기 분리 전과 FMT 양성 세포의 두 가지 하위 집합으로 나뉩니다. 유세포 분석은 12-48시간 후에 MDSC의 거의 60%와 대식세포의 60% 이상이 FMT를 축적하는 것으로 나타났습니다(그림 1b, c). DC의 약 40%만이 24 h에 FMT 양성이었습니다. 이러한 데이터는 대식세포와 마찬가지로 MDSC가 FMT를 흡수할 수 있음을 보여주고 FMT가 MDSC 기능에 영향을 미칠 수 있음을 시사했습니다.
<그림>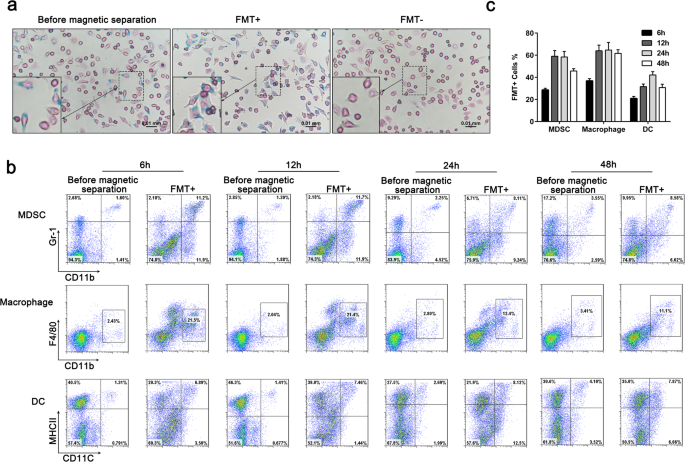
FMT를 흡수하는 MDSC, 대식세포 및 DC의 능력. 아 대식세포를 24시간 동안 FMT(1000 ng/mL)로 처리한 다음 Prussian blue로 염색하여 세 그룹 사이에서 철의 세포 존재 및 침착을 확인했습니다:자기 분리 전, 자기적으로 선택됨(FMT+), 자기적으로 분리되지 않음(FMT-) . ㄴ 나이브 C57BL/6 마우스의 비장세포를 6 h, 12 h, 24 h 및 48 h 동안 FMT와 함께 배양했습니다. 두 가지 하위 집합에서 MDSC, 대식세포 및 DC의 백분율에 대한 FACS 분석:자기 분리 전, FMT 양성 세포. ㄷ 자기 분리 전 세포에 대한 FMT 양성 세포의 비율. 모든 데이터는 각 실험 그룹에 대한 3개의 독립적인 실험을 대표하며 평균 ± 표준 편차로 표시됩니다.
FMT가 M2 대식세포에서 높은 CD86 + 쪽으로 표현형 이동을 유도하는 것으로 보고되었습니다. , TNF-α 양성 M1 대식세포 아형[3]. FMT를 사용할 수 있는 MDSC가 많기 때문에 FMT가 MDSC의 기능을 변경할 수 있다고 가정했습니다. 먼저, MDSC에 대한 250, 500, 1000 및 2000㎍/mL에서 FMT의 세포독성 효과를 CCK8 세포 생존력 분석에 의해 평가하였다. 결과는 FMT가 낮은 용량에서 세포 생존율에 영향을 미치지 않았으며 2000μg/mL의 최대 용량에서만 중간 정도의 세포 독성을 나타냈다는 것을 보여주었습니다(그림 2a). 그런 다음 다른 농도의 FMT가 MDSC의 생성에 영향을 미치는지 여부를 테스트했습니다. 나이브 C57BL/6 마우스에서 분리한 골수 세포를 배지 또는 다양한 농도의 FMT(250, 500, 1000 및 2000μg/mL)로 4일 동안 처리한 다음, 4일째에 유세포 분석으로 특성화했습니다. GM-CSF 및 IL-6은 0일에 추가되었습니다. 3개의 독립적인 실험 결과 1000 및 2000 μg/mL의 FMT가 MDSC의 확장을 유의하게 감소시키는 것으로 나타났습니다(그림 2b, c).
<그림>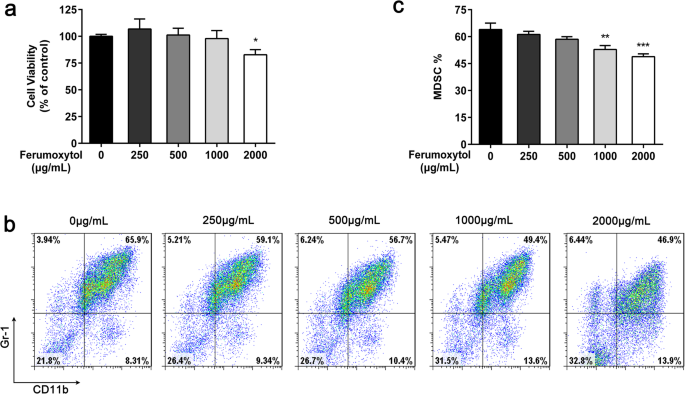
CD11b + 비율의 유세포 측정 정량화 1학년 + 시험관 내에서 250, 500, 1000, 2000 μg/mL의 FMT 처리 후 세포. 아 24시간 동안 다양한 농도의 FMT로 처리한 후 CCK8 분석을 사용하여 MDSC 세포 생존력을 검출했습니다. ㄴ , ㄷ C57BL/6 마우스로부터 분리된 골수 세포를 상이한 농도의 FMT 존재 하에 GM-CSF 및 IL-6과 함께 4일 동안 배양하고, 세포 표현형을 유세포 분석에 의해 평가하였다. 모든 데이터는 각 실험 그룹에 대한 3개의 독립적인 실험을 대표하며 평균 ± 표준 편차로 표시됩니다. *p <0.05, **p <0.01, ***p <0.001 짝을 이루지 않은 학생의 t에 의해 결정됨 시험 대 비히클 단독 처리
MDSC는 T 세포의 증식 및 효과기 기능을 억제하기 위해 여러 메커니즘을 사용하며, 가장 잘 설명된 것은 ROS 및 Arg-1의 생성입니다. nicotinamide adenine dinucleotide phosphate-oxidase (NOX) 복합체의 p47phox 성분 발현은 MDSC에서 ROS 생성을 담당합니다[5]. S100A8/A9는 자가분비 염증 루프에서 MDSC에 의해 합성 및 분비되며 이것이 마우스 모델에서 수지상 세포 기능의 억제에 중요하다고 보고되었습니다[16]. FMT가 MDSC의 기능을 변경할 수 있는지 확인하기 위해 우리는 이전에 설명한 대로 BM 유래 MDSC를 얻었고 RT-PCR을 사용하여 처리되지 않은 대조군과 비교하여 FMT 처리된 MDSC에서 mRNA 발현을 측정했습니다. 결과는 FMT 처리 MDSC가 처리되지 않은 MDSC와 비교하여 Arg-1, S100A8, S100A9 및 p47phox의 발현 수준을 유의하게 하향 조절하는 것으로 나타났습니다(그림 3a-h). 다음으로 대조군 MDSC와 FMT 처리 MDSC에서 ROS 수준을 평가했습니다. 결과는 FMT 처리 MDSC가 대조군 세포보다 유의하게 낮은 수준의 ROS를 나타냄을 보여주었습니다(그림 3i, j). 우리는 이후에 FMT 처리군과 처리되지 않은 군에서 MDSC 하위 집단의 변화를 비교했습니다. 결과는 FMT가 G-MDSC의 비율을 감소시켰고 M-MDSC의 약간의 증가도 관찰됨을 보여주었습니다(그림 3k, l).
<그림>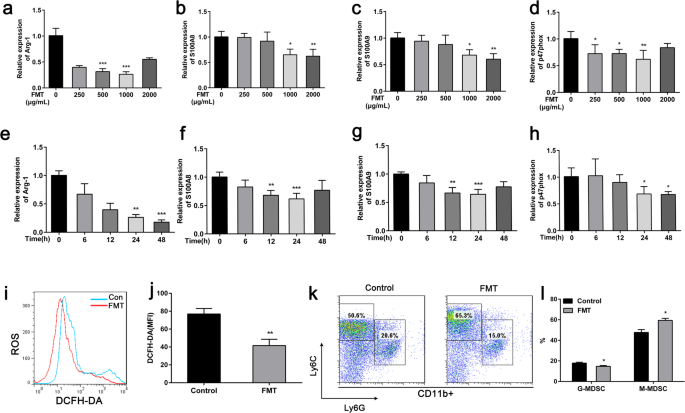
FMT는 시험관 내에서 MDSC의 기능을 변경합니다. 24 시간 동안 비히클 또는 FMT로 처리된 골수 유래 MDSC. 아 –h 정량적 RT-PCR로 측정한 처리된 MDSC에서 Arg-1, S100A8, S100A9 및 p47phox 유전자 발현. 나 BM 유래 MDSC를 FMT와 함께 24시간 동안 배양하고 FCM으로 ROS 생성을 검출했습니다. j i의 양적 데이터 . 케이 FMT 처리 및 대조군에서 MDSC의 하위 집단. 나 G-MDSC 및 M-MDSC의 백분율에 대한 FACS 분석. 모든 데이터는 각 실험 그룹에 대한 3개의 독립적인 실험을 대표하며 평균 ± 표준 편차로 표시됩니다. *p <0.05, **p <0.01, ***p <0.001 짝을 이루지 않은 학생의 t에 의해 결정됨 시험 대 비히클 단독 처리
FMT가 MDSC의 대식세포로의 최종 분화를 자극하는지 여부를 결정하기 위해 우리는 0일에 1000μg/mL FMT로 세포를 처리하고 1, 3, 5일에 대식세포 관련 마커 CD11b 및 F4/80에 대해 유세포 분석을 수행했습니다. 결과는 1000 μg/mL FMT를 첨가한 후 1일에 대식세포의 비율이 처리되지 않은 세포와 비교하여 2배 이상 크게 증가한 것으로 나타났습니다. 유사하게 FMT 처리 3일 및 5일 후 대식세포의 비율도 유의하게 증가하였다(Fig. 4a, b).
<그림>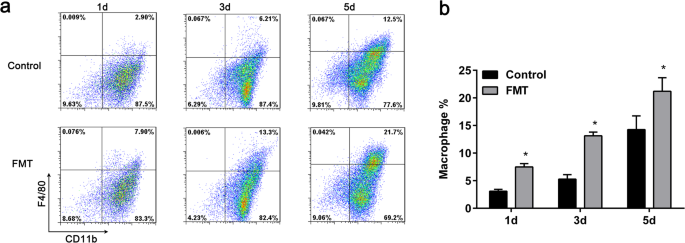
MDSC의 분화에 대한 FMT의 효과. 아 , b 골수 유래 MDSC는 1000 μg/mL FMT의 유무에 관계없이 처리되었고 대식세포의 수는 FACS에 의해 1일, 3일 및 5일 후에 평가되었습니다. 모든 데이터는 각 실험 그룹에 대한 3개의 독립적인 실험을 대표하며 평균 ± 표준 편차로 표시됩니다. *p <0.05, 비히클 단독으로 처리된 세포에 비해
패혈증은 조직 손상과 연관되어 기관 부전 및 내피 기능 장애를 유발하는 것으로 보고되었습니다[17]. FMT가 패혈증 후기 증상을 감소시키는지 알아보기 위해 10일 후 실험 쥐의 장기 손상 정도를 평가했습니다. LPS 유발 패혈증에서 폐의 조직병리학적 검사는 FMT가 비히클 처리된 마우스와 비교하여 백혈구 모집 및 폐포벽 비후를 감소시키는 것으로 나타났습니다. H&E 염색은 또한 간 괴사 영역이 비히클 처리된 마우스에 비해 FMT 처리된 마우스에서 더 작은 것으로 나타났습니다(그림 5a, b). 또한, 이들 동물에서 간 기능 장애의 생화학적 마커로 사용되는 아스파르테이트 트랜스아미나제 혈청 수준은 대조군과 비교하여 FMT 처리 마우스의 간에서 유의하게 낮았다(그림 5c). 그러나 FMT는 alanine aminotransferase의 수준에 영향을 미치지 않았습니다(그림 5d). 또한, 패혈증은 염증성 사이토카인의 생성 증가와 같은 부작용과 관련이 있는 것으로 생각됩니다. 따라서 우리는 혈청에서 전염증성 사이토카인 수준의 변화를 평가했습니다. 혈청 내 TNF-α의 수준은 LPS 주입 후 3 h에 FMT 처리된 생쥐에서 약간 낮았지만 유의하지는 않았으며, FMT 처리된 생쥐와 비히클 간에 MCP-1 또는 IL-1β 수준의 차이는 관찰되지 않았습니다. -처리된 마우스(그림 5e–g).
<그림>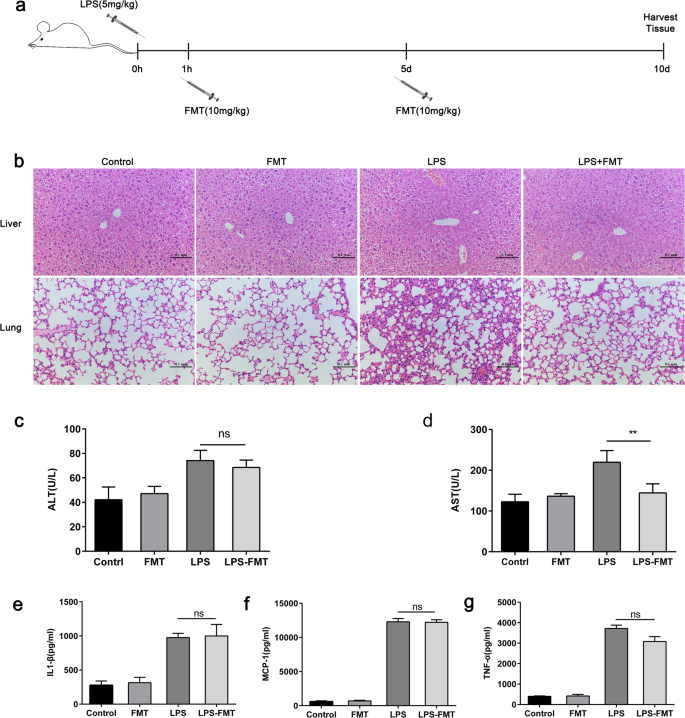
FMT는 LPS로 유도된 패혈증 마우스에서 간 손상을 개선합니다. 아 마우스는 PBS 또는 i.p. 0일째에 5 mg/kg 체중의 세균성 LPS 주사. 1 시간 및 5 일 후, FMT 10 mg/kg 또는 100 μL 식염수를 꼬리 정맥을 통해 투여했습니다. 마우스를 4개의 그룹(n =6):제어, 수신 차량; LPS, LPS만 수신 FMT, FMT만 수신 및 LPS + FMT, LPS 및 FMT 수신. ㄴ 폐와 간 조직은 헤마톡실린과 에오신으로 염색되었습니다. c-d 혈청 트랜스아미나제(ALT 및 AST)를 측정했습니다(n =6). 사이토카인 수준(TNF-a, IL-1β 및 MCP-1)을 ELISA로 측정했습니다. 이 -지 LPS 유도 패혈증이 비히클 또는 페루목시톨 처리 마우스에서 사이토카인 생성에 미치는 영향. i.p. LPS 챌린지(50 mg/kg). **p <0.01 짝을 이루지 않은 학생의 t에 의해 결정됨 테스트
패혈증 관련 면역억제가 사망률을 증가시키고[18], MDSC가 면역억제 네트워크의 주요 구성요소로 간주된다는 증거가 늘어나고 있습니다[8]. G-MDSCs는 패혈증 환자에서 보다 구체적으로 확장되며 패혈증 유발 면역 억제의 주요 행위자로 나타나는 것으로 보고되었습니다[12]. FMT가 야생형 마우스에서 MDSC 개체군을 조절하는지 여부를 확인하기 위해 C57BL/6 마우스에 1시간 및 5일에 꼬리 정맥을 통해 FMT를 투여했습니다. 결과는 MDSC의 비율에서 유의한 차이가 비장 또는 뼈에서 검출되지 않았다는 것을 보여주었습니다. 대조군과 FMT 처리 마우스 사이의 골수(그림 6a, e). 참고로 FMT는 CD11b + 의 LPS 종속 확장을 감소시켰습니다. 1학년 + 골수와 비장 모두의 세포(그림 6a, e)는 시험관 내 연구와 일치합니다. 또한 FMT는 비장에서 G-MDSC와 M-MDSC의 비율도 감소시켰지만 골수에서는 G-MDSC의 비율만 감소했습니다(그림 6c, g). 이 데이터는 FMT가 LPS로 유도된 패혈증 마우스의 비장에서 MDSC의 수를 감소시킨다는 것을 나타냅니다.
<그림>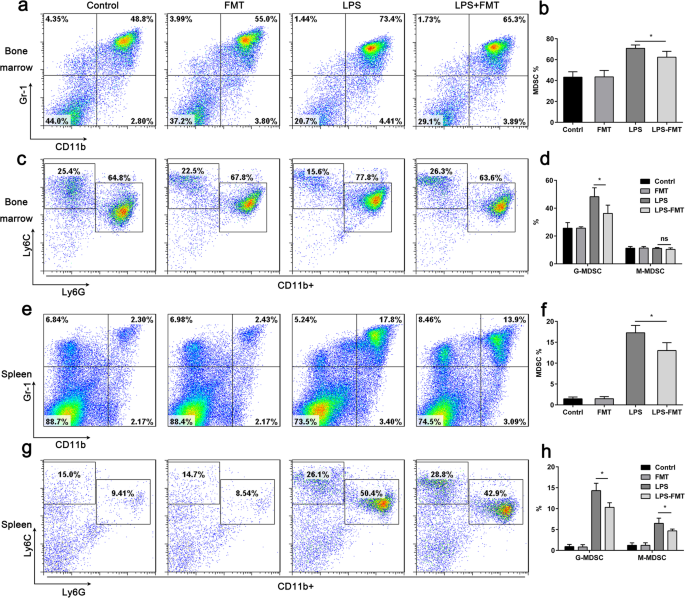
FMT는 LPS로 유도된 패혈증 마우스에서 MDSC의 수를 감소시킵니다. 마우스를 4개의 그룹(n =6):제어, 수신 차량; LPS, LPS만 수신 FMT, FMT만 수신 및 LPS + FMT, LPS 및 FMT 수신. 아 CD11b + 1학년 + 골수의 세포 집단은 FACS에 의해 검출되었습니다. ㄴ a의 양적 데이터 . ㄷ 게이트된 CD11b + 에서 Ly6C 및 Ly6G 발현의 유세포 분석 도트 플롯 비장 세포. d 과립구 및 단핵구 하위 집합의 백분율 정량화. 이 CD11b + 1학년 + 비장의 세포 집단은 FCM에 의해 검출되었습니다. 에 e의 양적 데이터 . 지 비장으로부터의 게이트된 CD11b+ 세포에서 Ly6C 및 Ly6G 발현의 유세포 분석 도트 플롯. 아 과립구 및 단핵구 하위 집합의 백분율 정량화. *p <0.05 짝을 이루지 않은 학생의 t에 의해 결정됨 테스트
CD4 및 CD8 T 세포를 포함한 다중 면역 세포 집단의 수치적 감소는 패혈증 후 적응 면역 반응의 주요 특징입니다. 사망 위험 증가 및 기타 좋지 않은 결과와 관련이 있습니다[19, 20]. MDSC는 패혈증 환자에서 T 세포 제타 사슬 발현을 손상시켜 T 세포 증식을 억제하는 것으로 나타났습니다[21]. 말기 패혈증에서 림프구의 비율을 평가하기 위해 쥐 패혈증 모델에서 CD4 및 CD8 T 세포의 비율을 측정했습니다. 결과는 FMT가 LPS로 유도된 마우스의 비장에서 CD4 및 CD8 T 세포의 증가를 유발함을 보여주었습니다(그림 7a, b). 대조군 마우스와 비교하여 FMT 처리 마우스의 비장에서 CD4 및 CD8 T 세포의 백분율에는 차이가 없었습니다(그림 7a, b). 생체 내에서 T 세포 증식을 억제하는 MDSC의 역할을 추가로 확인하기 위해, MACS에 의해 비히클 또는 FMT 처리된 LPS-챌린지 마우스의 비장에서 G-MDSC 및 M-MDSC를 분리하고 CD3 T 세포와 1:1 비율로 공배양했습니다. 비율. 우리의 결과는 FMT 처리가 G-MDSC와 M-MDSC 모두의 T 세포 면역억제 기능을 감소시켰음을 보여주었다(그림 7e, f). 또한, 우리는 또한 마우스의 비장에서 분리된 MDSC에서 Arg-1 및 p47phox 발현 수준을 측정했습니다. 예상대로 Arg-1 및 p47phox 유전자 발현은 비장 CD11b + 에서 크게 감소했습니다. 1학년 + LPS-챌린지 마우스와 비교할 때 FMT-처리 LPS-챌린지 마우스에서 분리된 세포(그림 7g, h). 이러한 데이터는 FMT가 Arg-1 및 ROS 생성을 감소시켜 MDSC의 면역억제 기능을 감소시켰음을 나타내며, 이는 FMT가 말기 패혈증에서 장기간 면역억제를 감소시킬 수 있음을 시사합니다.
<그림>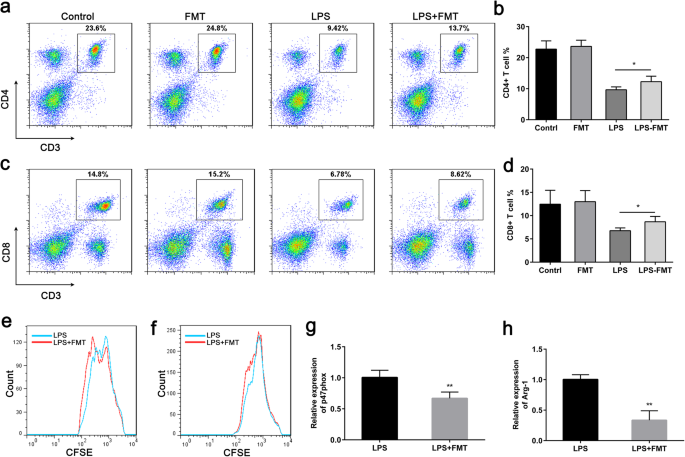
FMT는 MDSC의 면역 억제 특성을 감소시킵니다. 아 CD3 + CD4 + 비장의 세포 집단은 FACS에 의해 검출되었습니다. ㄴ a의 양적 데이터 . ㄷ CD3 + CD8 + 비장의 세포 집단은 FACS에 의해 검출되었습니다. d a의 양적 데이터 . 이 항-CD3/CD28 자극에 의해 유도된 T 세포 증식을 억제하는 마우스의 G-MDSC의 능력은 FACS에 의해 측정되었습니다 =그룹당 3개). 대표적인 히스토그램이 표시됩니다. 에 항-CD3/CD28 자극에 의해 유도된 T 세포 증식을 억제하는 비히클 또는 FMT 처리된 LPS-챌린지 마우스의 M-MDSC의 능력은 FACS에 의해 측정되었습니다(n =그룹당 3개). 대표적인 히스토그램이 표시됩니다. 지 , h 비히클 또는 FMT 처리 LPS 감염 마우스의 MDSC에서 Arg-1 및 p47phox mRNA 발현
FDA 승인 철분 보충제 FMT는 MRI 스캔에서 약물 운반체 및 조영제로 널리 사용되었습니다. FMT가 M1 대식세포 분극화로 전염증성 면역 반응을 유도하여 암 성장을 억제하는 것으로 밝혀졌으며[3], 이는 FMT가 면역 조절 기능을 가지고 있음을 나타냅니다. 이 연구에서 우리는 FMT 처리 MDSC의 표현형과 기능을 처리되지 않은 대조군 MDSC와 비교하여 MDSC에 대한 FMT의 효과를 평가했습니다. 이전 연구에서는 MDSC가 자성 나노입자를 흡수할 수 있음을 보여주었고 [10], 우리의 데이터는 FMT가 Arg-1 및 ROS의 하향 조절을 통해 비장에서 MDSC의 면역 억제 기능을 약화시킨다는 것을 추가로 보여주었습니다. 우리의 데이터는 또한 FMT가 시험관내 실험에서 5일 후에 대식세포로 고르게 분화하는 세포의 약 5분의 1과 함께 MDSC 분화에 직접적인 영향을 미치는 것으로 나타났습니다. FMT에 노출된 대식세포의 표현형을 조사하지 않았지만, 우리는 그들이 전염증성 M1 아형에 속한다고 가정했습니다. 결과는 FMT가 MDSC의 분화와 성숙을 개선한다는 것을 분명히 보여주었고, 따라서 이것이 후기 패혈증 동안 관찰된 MDSC 개체군 감소의 기초가 됨을 시사합니다.
패혈증은 시간이 지남에 따라 변하는 복잡한 면역 반응을 시작하며 전염증 및 항염증 반응을 동반합니다. 이전 연구에서는 패혈증 초기 단계의 사망이 과도한 염증으로 인한 것으로 나타났습니다. Derive와 동료들은 패혈증 마우스에 10일째 MDSC를 입양한 것이 복막 사이토카인 생성을 약화시키고 생존율을 향상시켰다고 보고했습니다[22]. 보다 최근에는 Namkoog et al. clarithromycin 전처리는 CD11b + 를 확장하여 LPS 유발 쇼크의 마우스 모델에서 생존을 향상시키는 것으로 관찰되었습니다. 1학년 + 세포 집단 [23]. They described that MDSCs play a protective role in sepsis, however, it should be noted that they were concerned about the early stages of sepsis. Most patients with sepsis display rapid signs of profound immunosuppression, and deaths in this phase are typically due to the acquisition of a secondary hospital-acquired infection, often with opportunistic pathogens [24]. Overzealous MDSC proliferation may facilitate a physiological syndrome of persistent immunosuppression, causing poor outcomes [25]. McClure et al. reported that they did not observe any protective effects from MDSC transfer once the mice entered the late immunosuppressive state [26]. They had previously shown that MDSCs massively expand in the bone marrow, spleen, and lymph nodes of mice with ongoing septic processes and contribute to sepsis-induced T cell suppression [11]. Additionally, Uhel reported that M-MDSCs and G-MDSCs strongly contribute to T cell dysfunction in patients with sepsis [12]. Our data demonstrated that FMT significantly decreased the percentage of MDSCs and attenuated the functions of MDSCs to restore the number of T cells present in mice during late sepsis. Therefore, when studying the role of MDSCs in sepsis, it is crucial to clearly distinguish the early and late stages of sepsis.
Mohus et al. suggested that iron deficiency was associated with increased risk of future bloodstream infections that cause sepsis, indicating that the immune defense mechanisms may be depressed compared to bacterial iron sequestration in low iron environments [27]. Controversies shown in iron supplemental studies reported that intravenous iron therapy was associated with an increased risk of infection [28]. It should be noted the iron supplements differ significantly in their physicochemical properties, which give them different biological properties [29]. With FMT, an iron core is wrapped in a carbohydrate shell, which leads to low toxicity, lysosomal uptake and degradation [30, 31]. It has been reported that FMT does not cause liver toxicity in patients or animal models and is typically metabolized within 2 months [32]. Our data showed that FMT does not increase the production of inflammatory cytokines in early sepsis, indicating that FMT can be administered in sepsis.
This study demonstrated a novel immune-modulatory property of FMT; however, further studies are needed to elucidate the mechanism of FMT suppression of MDSCs. Furthermore, we provide an attractive therapeutic approach for the treatment of sepsis-associated immunosuppression and targeting MDSCs may provide a promising new option for restoring the immune response during sepsis.
현재 연구 중에 사용 및/또는 분석된 데이터 세트는 합당한 요청이 있는 경우 교신 저자에게 제공됩니다.
알라닌 아미노전이효소
Arginase 1
Aspartate aminotransferase
Ferumoxytol
Polymorphonuclear MDSCs/granulocytic MDSCs
헤마톡실린-에오신
Interleukin-1β
Inducible nitric oxide synthase
지질다당류
Monocyte chemotactic protein 1
Myeloid-derived suppressor cell
Major histocompatibility complex
Monocytic MDSCs
Phosphate-buffered saline solution
활성 산소 종
Toll-like receptor
Tumor necrosis factor α
나노물질
출처:www. collaborativerobotarm.com 제조 산업에서 자동화 시스템의 사용은 빠르게 추진력을 얻고 있습니다. 이는 주로 이러한 자동화가 작업 출력 및 일반 성능에 가져다주는 편리함 때문입니다. 다양한 분야의 여러 제조업체에서 반복적인 작업과 고위험 작업을 처리하기 위해 로봇 팔의 사용을 수용하고 있습니다. 이 로봇을 사용하여 작업장에서 부상을 줄이고 직원이 다른 더 중요한 작업에 집중할 수 있습니다. 그러나 이러한 산업용 로봇 팔을 사용하려면 자유롭게 움직일 수 있도록 넓은 공간이 필요합니다. 작은 바닥
BJT 로드 라인 또는 Bipolar Junction Transistor는 전자와 전자 정공을 전하 캐리어로 제공합니다. 터미널 중 하나에 작은 전류를 주입할 수 있었습니다. 그런 다음 두 터미널 사이에 흐르는 더 큰 전류를 제어할 수 있습니다. 이 기능이 있는 장치는 신호를 증폭하거나 전환할 수 있습니다. BJT 부하 라인은 디지털 회로의 스위치 역할을 하는 트랜지스터입니다. 아날로그 회로에서도 증폭기로 작동합니다. 전반적으로 이 트랜지스터는 스위치를 켜고 끄는 데 도움이 됩니다. 아래에서 다양한 유형의 로드 라인과 그래프에