

나노물질

산업 제조
수많은 여성 생식기 이상이 자궁 평활근(자궁근) 장애로 인해 발생합니다. 중금속은 자궁 평활근의 수축력에 부정적인 영향을 미칩니다. 아연은 대부분의 유기체에 필수적인 생물학적 요소이지만 이 요소의 고용량은 유독합니다. 0.5-5mM Zn 2+ 연구 자궁에서 미오신 S1 ATPase 활성에 대한 효과는 5mM Zn 2+ 양이온이 가장 두드러진 억제 효과를 나타냅니다. 운동 매개변수의 계산(K m 및 V 최대 , ATP )는 5mM Zn 2+ 의 존재 하에 미오신에 의해 촉매되는 가수분해 ATP의 겉보기 최대 속도를 밝혔습니다. 1.6배 감소했습니다. К의 값 m Zn 2+ 존재하에서 미오신 S1에 의한 ATP 가수분해 감소하는 경향이 있지만 통계적으로 변하지 않습니다. 자궁 미오신 S1 ATPase 활성은 Mg 2+ 농도에 의존하지 않는 것으로 결정되었습니다. 5mM Zn 2+ 존재 시 . 또한, 테트라히드록시티아칼릭스[4]아렌-테트라술포네이트(C-798)와 테트라히드록시티아칼릭스[4]아렌-테트라포스포네이트(C-800)는 5mM Zn 존재하에서 미오신 S1 ATPase 활성을 대조군 수준으로 회복시키는 것으로 입증되었습니다. 2+ . 이러한 티아칼릭스[4]아렌 보호 효과의 작용을 회복하는 가장 가능성 있는 메커니즘 중 하나는 배양 배지에서 중금속 양이온을 킬레이트화하는 능력에 기반합니다. 미오신 S1 영역에 C-798과 C-800의 분자 도킹은 이들 티아칼릭스[4]아렌이 ATPase 활성 부위 근처의 미오신 아미노산 잔기에 의해 Zn 양이온 결합과 상호작용할 수 있음을 보여주었다. 따라서 thiacalix[4]arenes는 이 양이온과 myosin S1 사이의 상호작용을 약화시킬 수 있습니다. 얻어진 결과는 이 티아칼릭스[4]아렌을 고농도 아연 중독의 경우 약리학적 화합물로 사용하는 것을 목표로 하는 추가 연구에 사용될 수 있을 것으로 추측되었습니다.
<섹션 데이터-제목="배경">중금속으로 인한 환경 오염 문제와 중금속이 생물체에 미치는 영향을 줄이는 방법을 찾는 것은 관련이 있습니다[1, 2].
아연은 대부분의 유기체에 필수적인 생물학적 요소입니다. [삼]. 아연 이온은 중요한 대사 기능을 수행하는 여러 단백질과 복합체를 형성합니다. 아연 이온은 50가지 이상의 다양한 생화학적(생리학적) 반응을 촉매하는 300가지 이상의 금속 효소의 구성 요소입니다[4, 5].
그러나 아연은 중금속입니다. 그것은 두 가지 독성 금속 카드뮴 및 수은과 함께 원소 주기율표의 IIb 족에서 찾을 수 있습니다. 그럼에도 불구하고 아연은 인간에게 상대적으로 독성이 없는 것으로 간주됩니다[6]. 이 요소는 과다 복용 시에만 유해합니다[7].
구두 LD50 미국 국립 의학 도서관의 TOXNET 데이터베이스에 따르면 아연의 경우 체중 kg당 3g에 가깝습니다. 카드뮴보다 10배, 수은보다 50배 높다[6]. 인간에서이 미량 원소의 정상 농도의 초과는 가장 일반적으로 구성에 중복 아연을 포함하는 약물 및 생물학적 활성 첨가제의 섭취로 인해 발생합니다. 아연 코팅 또는 완전 아연 용기에 저장된 음식을 섭취한 결과 아연 중독의 개별 사례로 기록되었습니다. 산화아연, 염화물 및 황산아연은 업계에서 광범위하게 사용됩니다. 유리 생산을 위해; 인공 섬유, 아연 페인트, 도자기, 성냥 및 치과용 시멘트 제조; 펄프 및 제지 산업, 목재 보존, 주석 도금 및 납땜용
고농도의 아연 섭취는 면역 반응을 변화시켰습니다[8]. 일부 역학 연구에 따르면 뇌의 Zn, Al, Cu 및 Fe 수치가 높아지면 알츠하이머 병의 발병 또는 진행이 촉진될 수 있다고 합니다[9, 10].
중금속은 태아 생활의 시작, 초기 발달 및 성숙과 같은 여러 단계에서 여성의 생식에 영향을 줄 수 있습니다. 중금속 양이온은 또한 불임, 불임, 자궁 내 성장 지연, 자연 유산, 기형, 기형, 출생 후 사망, 조기 노화, 학습 및 행동 장애의 원인이 될 수 있습니다[11, 12].
자궁 수축 기능은 미오신이 효소 활성, 즉 ATP를 가수분해하는 능력을 나타내는 단백질 복합체(액토미오신)의 활성과 관련이 있습니다. subfragment-1(S1 또는 헤드)의 촉매 도메인에 국한된 미오신 ATPase는 ATP의 거대 결합에 축적된 화학 에너지를 기계적 운동으로 변환합니다. 결과적으로 미오신은 액틴 필라멘트를 따라 이동하여 근육 수축을 유발합니다. 따라서 myosin에 의해 촉매되는 ATP 가수분해는 myometrial 기능의 분자적 기전에서 필수적인 과정 중 하나로 간주된다[13, 14].
미오신 하위 단편-1은 ATP-ase 부위와 액틴 결합 부위를 포함하는 N-말단 구형 운동(촉매) 도메인과 조절 도메인의 두 도메인으로 구성된 미오신 중쇄의 N-말단 부분입니다. 또는 액틴 필라멘트를 따라 미오신의 움직임을 담당하는 레버 암. 미오신 모터 도메인의 핵심은 α-나선으로 둘러싸인 중앙의 7가닥 β-시트에 의해 형성됩니다. 중앙 β-시트의 7개 가닥 중 6개를 차지하는 큰 구조 도메인은 일반적으로 상위 50kDa 도메인(U50)이라고 합니다. 큰 틈은 465에서 590까지의 아미노산 잔기에 의해 형성되는 잘 정의된 구조적 하부 50kDa 도메인(L50)으로부터 상위 50kDa 도메인을 분리합니다. 미오신의 액틴 결합 영역과 뉴클레오티드 결합 부위는 7-가닥 β-시트의 반대쪽에는 뉴클레오티드 결합 포켓의 뒤쪽에 뉴클레오티드의 인산염 모이어티가 있습니다. P 루프, 스위치 1 및 스위치 2는 큰 틈새의 정점에 가까운 상위 50kDa 영역에 있습니다. 3개의 뉴클레오티드 결합 모티프는 모두 뉴클레오티드 결합 포켓 뒤쪽에 있는 뉴클레오티드의 인산염 부분과 접촉하고 γ-인산염 센서로 작용합니다[15].
우리의 이전 연구에서 중금속 양이온이 자궁 평활근의 미오신 ATPase 활성을 억제한다는 것이 밝혀졌습니다[16, 17]. 이는 자궁근의 수축 특성에 부정적인 영향을 미칠 수 있습니다.
자궁 수축에 대한 중금속의 역효과는 이러한 유해한 영향을 제거할 수 있는 약리학적 물질의 개발이 필요합니다.
Calixarene은 현재 다양한 생화학적 과정을 위한 인공 이펙터로 연구자들의 관심을 끌고 있습니다. 이 화합물은 컵 모양의 구조를 갖는 합성 거대고리 페놀 올리고머입니다. 상부 및 하부 테두리는 다양한 화학적 치환기로 기능화될 수 있습니다. Calix[4]아렌은 4개의 기능화된 아렌 단편으로 형성되며 다소 유연한 거대고리 형태를 특징으로 합니다. Calix[4]arenes는 기질의 낮은 독성과 세포 침투 능력을 보였다. 따라서 이들 화합물은 새로운 효과적인 약물 개발을 위한 유망한 약제로 간주됩니다[18, 19].
그러한 물질의 유망한 부류는 거대고리 분자 플랫폼에서 금속 착물기를 갖는 수용성 티아칼릭사렌이다[18]. 칼릭사렌은 (생체)금속 양이온과 초분자 복합체를 형성하는 능력으로 인해 중금속 추출제로 생물의학 연구에서도 사용되었습니다[20,21,22].
우리는 이전에 tetrahydroxy-thiacalix[4]arene-tetrasulfonate(С-798)가 Pb 2+ 의 억제 효과를 제거했음을 보여주었습니다. , CD 2+ , Ni 2+ 돼지 자궁의 미오신 S1에 의해 촉매되는 ATP 가수분해 [23].
이 연구는 고농도 아연 양이온과 tetrahydroxy-thiacalix[4]arene-tetrasulfonate (С-798) 및 tetrahydroxy-thiacalix[4]arene-tetraphosphonate (C-800)와의 결합 작용이 미오신에 미치는 영향을 연구하는 것을 목적으로 합니다. 자궁에서 S1 ATPase 활동. 이 연구는 자궁 미오신의 효소 활성에 대한 고농도 아연의 부작용을 제거하는 이러한 티아칼릭사렌의 능력을 테스트하는 데 필요했습니다.
Thiacalix[4]arene C-798 및 C-800은 각각 4개의 음이온성 설포네이트 및 4개의 포스포네이트 기로 상부 가장자리에서 변형된 4개의 페놀성 단편으로 형성된 컵으로 구성됩니다. 티아칼릭스[4]아렌은 모두 수산기와 2가 황 원자가 아래쪽 가장자리에 조밀하게 위치하여 안정적인 금속 착물을 형성하여 중금속을 킬레이트화할 수 있습니다[21](그림 1).
<사진>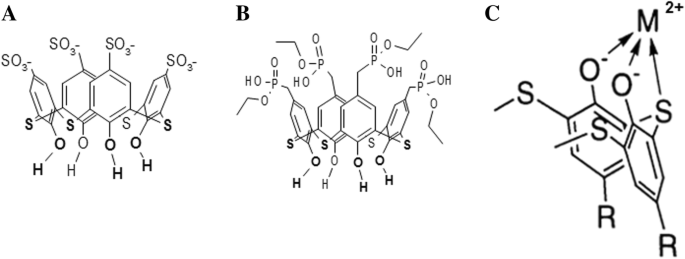
테트라하이드록시-티아칼릭스[4]아렌-테트라설포네이트(C-798)의 화학 구조(a ), 테트라하이드록시-티아칼릭스[4]아렌-테트라포스포네이트(C-800)(b ), 그리고 아래쪽 테두리에 금속 양이온이 있는 티아칼릭사렌의 킬레이트화 착물의 계획(역위 위치)(c )
이 연구는 자궁근층의 myosin ATPase와 자궁 myosin ATPase의 억제제 또는 활성화제(이펙터)인 calix[4]아렌의 상호작용에 초점을 맞춘 우크라이나의 Palladin Institute of Biochemistry and Institute of Organic Chemistry의 공동 프로젝트의 결과입니다.
<섹션 데이터-제목="결과">자궁에서 미오신 S1 ATPase 활성에 대한 가장 두드러진 억제 효과는 Zn 양이온의 경우 5mM(43 ± 8%, M ± SD)인 것으로 나타났습니다. Zn 2+ 의 농도 범위 배양 배지에서 0.5-5mM이었습니다(3mM ATP, 5mM Mg 2+ 포함) 및 0.01mM Ca 2+ ). 100%는 Zn 양이온(대조군)을 추가하지 않은 ATPase 활성 값입니다(그림 2). 따라서 미오신 S1 ATP 가수분해에 대한 Zn 양이온의 역효과는 5mM의 Zn 2+ 을 사용하여 추가로 연구되었습니다. .
<그림>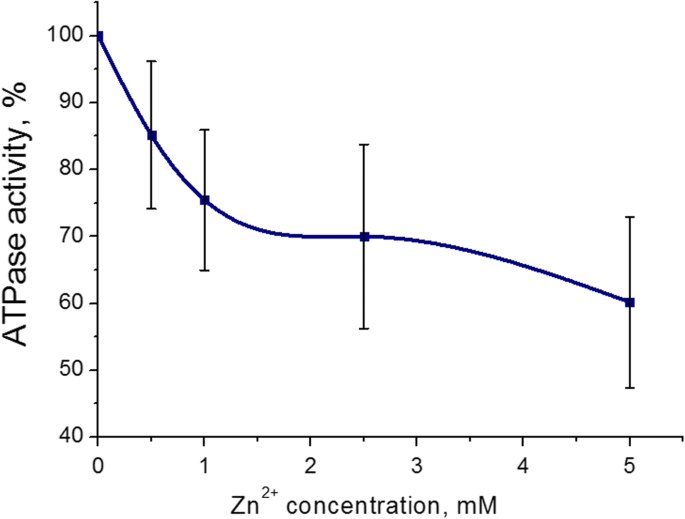
0.5–5.0mM 농도의 Zn 양이온(M ± SD, n)이 있는 경우 자궁근층에서 미오신 S1 ATPase 활성 =6). 100%는 Zn 양이온을 추가하지 않은 ATPase 활성 값입니다.
Zn 2+ 기질(ATP)에 대한 미오신 S1 ATPase 활성의 친화도에 대한 효과를 조사했습니다. 고정 MgCl2에서 배양 배지의 ATP 농도를 0.5에서 5mM으로 증가 대조군 및 5mM Zn 2+ 존재 시 농도(5mM) 그 결과 3mM ATP에서 ATPase 활성의 최대값이 있는 돔 모양의 그래프가 생성되었습니다. 아연이 존재할 때 이 피크의 효소 활성 값은 대조군보다 30% 낮았습니다(그림 3). 대조군의 ATP 농도와 5mM Zn 2+ 의 존재에 대한 미오신 S1 ATPase 활성의 의존성 그래프 Lineweaver-Burk 방법[27]에 따라 오름차순 부분에서 선형화되었습니다. 운동 매개변수의 계산, 즉 Michaelis의 허수 상수(K m ) 및 ATP에 대한 미오신 S1 ATPase의 최대 속도(V 최대, ATP )는 V 최대, ATP 5mM Zn 2+ 존재 시 미오신 효소 활성 1.6배 감소(대조군 및 Zn 2+ 의 존재에서 1 mg 단백질당 38 ± 7 및 22 ± 6 nmol Pi/min) 각각 n =5). К의 값 m 미오신 S1에 의한 ATP 가수분해의 경우 통계적으로 변화하지 않지만 감소하는 경향이 있습니다(대조군에서 0.49 ± 0.15mM, Zn 2+ 존재 시 0.38 ± 0.12mM); M ± SD; n =5).
<사진>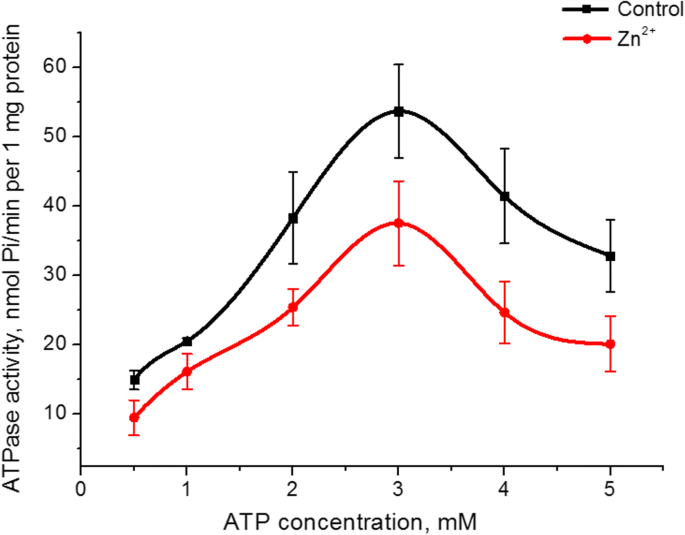
5mM Zn 2+ 이 있는 경우 자궁의 미오신 S1 ATPase 활성에 대한 0.5~5mM ATP의 영향 대조군과 비교(M ± SD, n =5)
5mM Zn 2+ 의 효과 Mg 2+ 에서 자궁 미오신의 ATPase 활성에 대한 농도 의존성을 연구했습니다. Mg 증가 2+ 5mM Zn 2+ 존재 하에 고정 ATP 농도(3mM)에서 0.5~5mM의 배양 배지 농도 미오신 S1 ATPase 활성의 변화를 일으키지 않습니다. 동시에 Mg 2+ 대조군(표준 조건에서)에서 ATPase 활성의 농도 의존성이 검출되었습니다. 대조군에서 가장 높은 수준의 미오신 ATP 가수분해는 3mM Mg 2+ 에서 달성되었습니다. (그림 4). 따라서 자궁의 미오신 S1 효소 활성은 Mg 2+ 농도에 의존하지 않습니다. Zn 고농도(5mM)가 있는 경우.
<그림>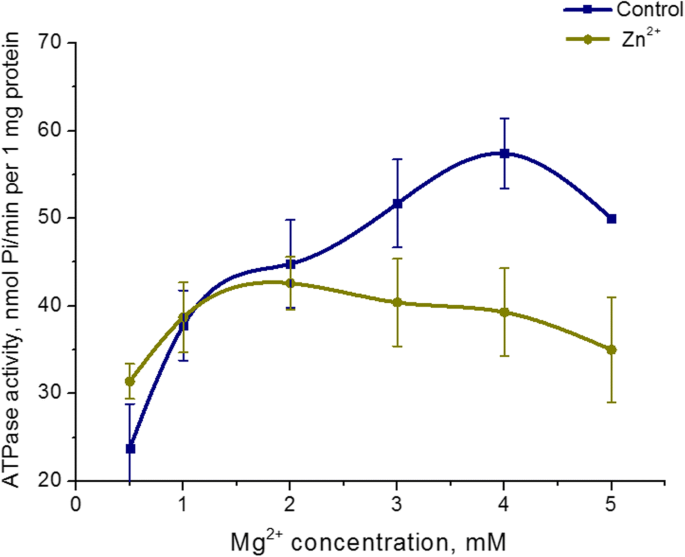
Mg 2+ 에 대한 미오신 S1 ATPase 활성 의존성 5mM Zn 2+ 존재 시 농도 대조군과 비교(M ± SD, n =6)
컴퓨터 시뮬레이션은 Zn 양이온이 미오신 머리에 여러 결합 영역을 가지고 있음을 보여줍니다. 그 중 하나는 뉴클레오티드 결합 부위에 가깝고 P 루프 바로 근처에 있는 상위 및 하위 50kDa 하위 도메인 사이의 틈의 아래쪽 부분에 있습니다. Zn 2+ 산소 원자 Glu177(결합 길이 0.23 및 0.39nm), Ser178 산소 원자(결합 길이 0.31nm) 및 Arg236(결합 길이 0.32nm)과 배위됩니다.
다른 Zn 결합 영역은 상위 50kDa 하위 도메인(Leu218-Asp463, Glu605-Phe621)의 하단에 있으며 스위치 1(Gly233-Phe246) 및 P-루프에 가깝습니다. Zn 2+ Glu327 산소 원자(결합 길이 0.21nm), 산소 원자 Glu326(결합 길이 0.34nm) 및 산소 원자 Asp323(결합 길이 0.32nm)과 배위할 수 있습니다. Zn 양이온은 또한 스위치 2와 접촉하는 영역에서 미오신 S1과 상호작용하여 Glu 465(0.24nm), Asp468(결합 길이 0.31nm) 및 Leu653(결합 길이 0.37nm)과 상호작용할 수 있습니다. 이 결합 영역은 액틴 결합 부위 근처에 있으며 상위 50kDa 하위 도메인과 하위 도메인 사이의 틈입니다. 이 틈의 바닥은 ATP 결합 주머니에 있습니다. 이러한 결합 Zn 2+ 미오신 S1 도메인은 ATP의 결합 및 가수분해에 필수적인 역할을 합니다. 이 영역은 ATP 가수분해 부위에서 액틴 결합 표면으로 에너지를 전달하는 과정에서 복잡한 형태 변형을 겪습니다.
50mM Tris-HCl 완충액(pH 7.2)에 100마이크로몰의 C-800 또는 C-798 용액을 5mM Zn 양이온이 포함된 배양 배지에 첨가하여 Zn 2+ 의 부정적인 영향을 제거했습니다. 자궁 평활근 미오신 S1의 ATPase 활성. 대조군으로 배양 배지에 아연 및/또는 티아칼릭스[4]아렌을 첨가하지 않고 효소 활성으로 사용하였다. 화합물 C-800은 자궁근층의 미오신 S1의 ATPase 활성에 영향을 미치지 않는 것으로 나타났다(그림 5). 비록 화합물 C-798은 일정량의 Mg 2+ 추출과 가장 관련이 있는 미오신 S1 ATPase 활성에 대한 약간의 억제 효과를 나타내지만 [21], 인큐베이션 배지에서 활성 센터의 ATP 결합 및 가수분해에 필수적입니다. 그럼에도 불구하고 100μM C-798과 C-800은 5mM Zn 2+ 의 억제 효과를 제거했습니다. 미오신 S1에 의해 촉매되는 ATP 가수분해 과정.
<그림>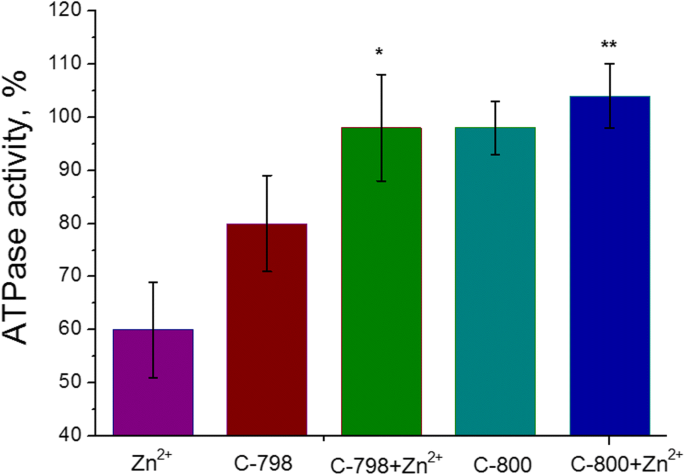
5mM Zn 2+ 존재 시 미오신 S1 ATPase 활성에 대한 100μM C-798 및 C-800의 효과 (M ± SD, n =5–6). 100%는 Zn 양이온을 첨가하지 않은 ATPase 활성 값입니다. "Zn"과 "Zn + C-798"의 차이, "Zn"과 "Zn + C-800"의 값 사이의 차이는 통계적으로 유의합니다(p <0.05) 및 각각 * 및 **로 표시
Zn 2+ 에서 미오신 S1 ATPase 활성에 대한 C-798 및 C-800 회복 작용의 가장 가능성 있는 메커니즘 중 하나 존재는 Zn 양이온에 결합하여 결과적으로 배양 배지에서 이러한 양이온을 배제하는 thiacalix[4]아렌의 능력일 수 있습니다. 이러한 티아칼릭스[4]아렌이 이미 미오신에 결합된 아연 양이온에 결합할 수 있는지 여부는 흥미로웠습니다.
컴퓨터 시뮬레이션은 방향족 고리 사이에 가교 황 원자가 있는 티아칼릭스[4]아렌 C-798 및 C-800이 페놀 그룹 사이의 수소 내 결합에 의해 안정화된 "원뿔" 형태임을 보여주었습니다. 이러한 calixes[4]아렌의 에너지 최소화 구조가 얻어졌습니다. 최소화 후 C-798의 총 에너지는 64.5kcal/mol이었다. 칼릭스[4]아렌 테두리(특히 아래쪽)에 이온화된 그룹이 있으면 호스트-게스트 상호 작용의 총 에너지에 대한 정전기 상호 작용의 기여도가 크게 증가합니다. 우리는 또한 C-798-Zn 2+ 의 "최소화"를 수행했습니다. 복잡한; 총 에너지는 83kcal/mole이었습니다.
C-798은 루프 P의 영역에서 단백질에 미리 결합되어 있던 Zn 양이온과 협력하는 미오신 S1의 구조에 박혀 있었다. 이 경우 Zn 2+ C-798(O3, 0.21nm, S1, 0.30nm, O2, 0.34nm)의 하부 테두리 산소 원자 및 브리지 황과 상호작용합니다. Zn 2+ P-loop의 아미노산 잔기에서 어느 정도 벗어나 Glu177 산소 원자(0.43nm 결합 길이)와의 상호 작용을 약화시킵니다(그림 6).
<그림>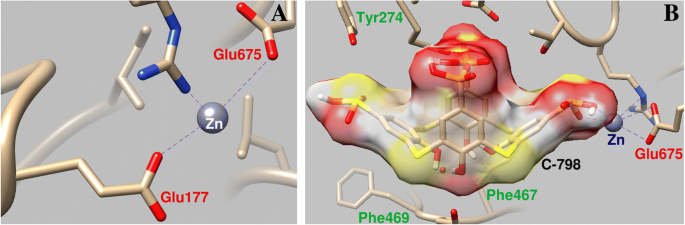
Zn 2+ 상호작용의 기하학적 매개변수 미오신 S1의 P 루프 영역(a ) 및 이 영역과 Zn 양이온의 협력에 대한 C-798의 영향(b )
미오신의 ATP 결합 영역의 "공동"에서 C-798의 고정은 여러 아미노산 잔기의 참여로 발생합니다. 특히, 티아칼릭사렌의 소수성 바스켓은 Phe467 및 Phe469의 미오신 방향족 아미노산 잔기에 의해 고정되고; 티아칼릭사렌은 음으로 하전된 산소 원자가 Arg570, Asn572 및 His689의 양으로 하전된 아미노산 잔기와 상호작용합니다.
Zn 2+ 의 변화에 대한 C-798의 영향 연구 미오신 ATP 결합 부위 근처 영역에 도킹하는 동안 위치는 Zn 2+ thiacalix[4]arene의 존재 하에서 세 번째 sulfonyl 그룹의 산소 원자(O16–0.26 nm, O15–0.27 nm)와 상호 작용하고, 산소 원자 Asp134 및 Glu326과 거의 상호 작용하지 않으며, Glu327과의 배위는 훨씬 약합니다. (0.43nm 커플링 길이)(그림 7). 이 경우 티아칼릭사렌은 여러 아미노산 잔기가 참여하여 단백질의 "공동"에 고정됩니다. 특히 티아칼릭사렌 설포닐기의 음전하 산소 원자는 Lys188, Lys195 및 Gln221의 미오신 양전하 아미노산 잔기와 상호작용합니다.
<사진>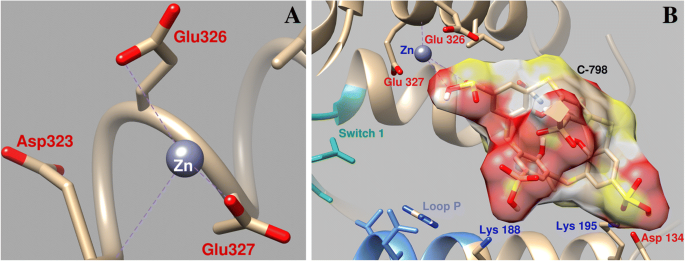
Zn 2+ 상호작용의 기하학적 매개변수 미오신 스위치 1과 미오신 S1의 P 루프(a ) 및 Zn 2+ 의 협력에 대한 C-798의 영향 이 지역(b )
Zn 양이온의 조건부 에너지 영역은 C-798의 산소 원자와 황 원자의 존재로 인해 정전기 상호 작용의 확산 표면과 접촉하고 심지어 다소 겹침을 보여줍니다. 이것은 연구 중인 양이온이 상부 및 하부 C-798 크라운의 음으로 하전된 원자와 충분히 밀접하게 상호작용하고 있음을 나타냅니다. 티아칼릭사렌이 스스로 Zn 2+ , 그 결과 효소의 아미노산 잔기와 양이온의 상호 작용이 약화됩니다.
결과적으로 Zn 2+ 을 포함하는 미오신 S1 영역에 C-798 도킹 결과 바인딩된 C-798 기능 그룹과 Zn 양이온의 상호 작용 가능성을 나타냅니다. 이 경우, 미오신 S1의 아미노산 잔기와의 Zn 양이온 결합이 현저히 약해지고, 이들 사이의 거리가 증가한다. 결과적으로 미오신의 ATPase 활성에 대한 Zn 양이온의 역효과를 제거할 수 있습니다.
우리는 또한 양이온 Zn의 기하학을 변경하기 위해 S1 미오신의 ATP 결합 영역에 가까운 영역에 도킹하여 calix[4]arene C-800의 효과에 대한 컴퓨터 시뮬레이션을 수행했습니다. 동시에 Zn 2+ Arg236(0.37nm 결합 길이), Glu675(0.41nm 결합 길이) 및 상부 기능 calix[4]아렌 잔기(H26, 2.36nm, H30, 2.96nm, C30, 0.31nm, O5, 3.7nm, O16, 0.4nm, O7, 0.48nm, O11, 0.51nm 결합 길이). Zn 2+ C-798과 유사하게 calix[4]arene C-800과 상호작용합니다. 양이온은 또한 이 영역의 이전 결합 부위에서 물러나며 Asp468(0.2nm 결합 길이) 및 하부 칼릭사렌 크라운의 원자(C4, 1.96nm, C14, 2.07nm, S3, 2.16nm, O2)와 접촉합니다. , 2.26nm, C3, 2.97nm, C20, 3.10nm, O4, 4.2nm, C14, 3.0nm, O3, 4.1nm)
미오신 하위 단편-1 영역에 도킹된 C-798 및 C-800은 이러한 티아칼릭스[4]아렌이 ATPase 활성 부위 근처의 미오신 아미노산 잔기에 결합하는 Zn 양이온과 상호작용할 수 있음을 보여주었습니다. 따라서 그들의 보호 효과는 이러한 양이온과 미오신 S1 사이의 상호 작용이 약해진 결과일 수 있습니다.
수많은 여성 생식기 이상이 자궁 평활근(자궁근) 장애로 인해 발생합니다. 중금속은 자궁 평활근의 수축력에 부정적인 영향을 미칩니다. 중금속 아연은 대부분의 유기체에 필수적인 생물학적 요소입니다. 이 요소의 고용량은 독성이 있습니다[5]. Zn 2+ 의 밀리몰 농도에 대한 여러 조사 살아있는 물체에 대한 것은 이전에 설명되었습니다[32]. 우리는 5mM Zn이 자궁의 미오신 S1 ATPase 활성에 가장 현저한 억제 효과를 갖는다는 것을 발견했습니다. 따라서 미오신 S1 ATP 가수분해에 대한 Zn 양이온의 역효과는 이 농도의 Zn 2+ 에서 더 연구되었습니다. . ATP에 대한 미오신 S1 ATPase의 운동 매개변수 계산은 V 최대, ATP 5mM Zn 2+ 존재 시 미오신 효소 활성 1.6배 감소했습니다. К의 값 m ATP 가수분해는 감소하는 경향이 있지만 통계적으로 변하지 않습니다.
생리학적 조건에서 미오신은 Mg 2+ 입니다. -의존성 ATPase. 마그네슘 양이온은 미오신 활성 부위의 ATP 결합과 ATP 가수분해에 관여합니다. Mg 2+ 효소의 활성 부위에서 미오신 아미노산 잔기 Thr-186과 Ser-237, 그리고 β-와 γ-의 형성과 함께 ATP 분자의 β- 및 γ-포스페이트 그룹과 함께 조정됩니다. bidentate complex와 활성 물 분자 중 하나가 γ-phosphate ATP에 대한 친핵체 공격을 수행합니다[33, 34]. 마그네슘 2+ ATP 음전하를 띤 인 그룹과 상호작용하여 극성화하여 말단 γ-포스페이트에 대한 친핵성 공격을 촉진합니다[14].
미오신 S1 ATPase 활성은 Mg 2+ 의 존재에 민감하지 않은 것으로 밝혀졌습니다. 5mM 농도의 Zn 2+ 에서 배양 배지에 아연이 없는 대조군과 대조적으로 [35, 36].
미오신 ATPase 활성은 금속 양이온 특성에 따라 달라지며 이온 반경과 잘 관련됩니다. 이온 반경 Mg 2+ 및 Zn 2+ 솔루션에서 매우 유사합니다(각각 0.070 및 0.076 nm)[37]. 따라서 Zn 2+ 의 상호작용 Mg 2+ 와 양이온 - 미오신의 결합 부위가 가능합니다. 따라서 Mg 2+ -결합 사이트는 Zn 2+ 이 차지할 수 있습니다. 높은 농도의 양이온. 이러한 조건에서 미오신 S1의 ATPase 활성은 마그네슘 양이온에 둔감할 수 있습니다. 미오신에는 Mg 2+ 에 대한 2개의 고친화성 사이트가 포함되어 있습니다. 및 Mg 2+ 이 부위에 결합된 근육은 근육 수축 동안 에너지 변환 과정에서 중요한 생리학적 역할을 합니다. 아직 여러 Mg 2+ 가 있습니다. - 미오신 분자의 ATPase 부위 외에 결합 부위로 마그네슘 이온의 결합 에너지와 친화도가 다릅니다[35]. 따라서 Zn 2+ ATP의 결합 및 가수분해에 영향을 미치는 미오신 S1의 다른 기능적으로 중요한 부위에도 결합할 수 있습니다.
컴퓨터 시뮬레이션은 Zn 양이온이 ATP 결합 부위, 즉 P-루프와 상위 및 하위 50kDa 하위 도메인 스위치 2에 가까운 미오신 헤드에 여러 결합 영역을 가지고 있음을 보여줍니다. 이러한 결합 Zn 2+ 미오신 S1 도메인은 ATP의 결합 및 가수분해에 필수적인 역할을 합니다. 이 영역은 ATP 가수분해 부위에서 액틴 결합 표면으로 에너지를 전달하는 과정에서 복잡한 형태 변형을 겪습니다.
Zn 2+ 로 얻은 분석 결과 미오신 S1에 도킹하는 것은 미오신 분자에 대한 이 양이온의 결합에서 핵심 역할이 효소 아미노산 잔기, 특히 Glu 및 Asp의 음으로 하전된 그룹과의 상호작용을 한다는 것을 나타냅니다.
Zn 2+ 의 독성 농도의 유해한 영향 미오신 S1 ATPase 활성에 대한 양이온은 이 금속의 작용을 제거할 수 있는 약리학적 화합물에 대한 검색을 필요로 합니다. 우리 연구의 대상은 안정한 금속 착물을 형성하여 전이 및 중금속을 킬레이트화할 수 있는 테트라히드록시티아칼릭스[4]아렌-테트라술포네이트(C-798) 및 테트라히드록시티아칼릭스[4]아렌-테트라포스포네이트(C-800)였습니다. 1). C-798과 C-800의 상부 거대고리형 크라운은 각각 4개의 음이온성 설포네이트기 또는 4개의 포스포네이트를 함유하고 있어 티아칼릭사렌의 우수한 수용성과 아미노산 단편의 양전하를 띤 질소 원자와의 정전기적 접촉으로 인해 단백질 분자에 대한 접착력을 제공합니다[21 ].
미오신 S1 영역에 도킹된 C-798 및 C-800은 이들 티아칼릭스[4]아렌이 ATPase 활성 부위 근처의 미오신 아미노산 잔기에 결합하는 Zn 양이온과 상호작용할 수 있음을 보여주었다. 따라서 그들의 보호 효과는 이러한 양이온과 미오신 S1 사이의 상호 작용이 약화 된 결과 일 수 있습니다. 얻어진 결과는 이 티아칼릭스[4]아렌을 고농도 아연 중독의 경우 약리학적 화합물로 사용하는 것을 목표로 하는 추가 연구에 사용될 수 있을 것으로 추측되었습니다.
고농도(5mM)의 Zn 양이온은 자궁에서 미오신 S1 ATPase 활성을 억제했습니다. Zn의 억제된 영향은 5mM Zn 2+ 존재 시 미오신 S1에 의해 촉매되는 가수분해 ATP의 최대 속도 감소와 관련이 있습니다. . К의 값 m ATP는 감소하는 경향이 있지만 통계적으로 변하지 않습니다.
Tetrahydroxythiacalix[4]arene-tetrasulfosphonate(C-798) 및 tetrahydroxythiacalix[4]arene-tetraphosphonate(C-800)는 5mM Zn 2+ 의 존재 하에서 미오신 S1 ATPase 활성을 대조군 수준으로 회복시켰습니다. .
Zn 양이온은 ATPase 활성 부위에 가까운 미오신 S1에 여러 결합 영역이 있습니다. Zn 2+ 을 포함하는 미오신 S1 영역에 C-798 및 C-800 도킹 결합은 이러한 티아칼릭스[4]아렌 작용기와 결합된 Zn 양이온의 상호작용 가능성을 나타냅니다. 미오신 S1의 아미노산 잔기와의 Zn 양이온 결합이 현저히 약해지고, 이들 사이의 거리가 증가한다. 결과적으로 미오신의 ATPase 활성에 대한 Zn 양이온의 역효과를 제거할 수 있습니다.
얻어진 결과는 아연 농도가 높은 중독의 경우 이 티아칼릭스[4]아렌을 약리학적 화합물로 사용하는 것을 목표로 하는 추가 연구에 사용될 수 있다고 가정합니다.
다음 시약이 사용되었습니다:혈청 알부민, EGTA, EDTA, ATP, 아스코르브산, Tris, 트리신, 디티오트레이톨, 아크릴아미드, (Sigma, USA), 글리신(Merck, Germany), N, N'-methylenebisacrylamide(Acros Organics, Belgium) ) N,N,N',N'-테트라메틸렌디아민(Reanal, 헝가리) 및 국내 생산 시약(R 등급). 용액은 Crystal Bio 시스템(라트비아의 Adrona)에서 정제된 물에서 제조되었습니다. 물 전도율은 0.1μS 미만이었습니다. 용액에서 2가 금속 양이온의 농도는 Mohr 방법에 의해 결정되었습니다.
Actomyosin was isolated from pig uterine smooth muscle by the modified Barany method as described in [17]. Myosin S1 was obtained from pig actomyosin by the modified Suzuki method [24]. The purity of the samples was controlled by PAAG-SDS electrophoresis [25].
ATPase activity of myosin S1 was determined in a 96-well plate at 37 °C in an incubation medium (total volume 0.1 ml) of the following composition (mM):Tris-HCl buffer (pH 7.2), 20; KCl, 100; CaCl2 , 0.01; MgCl2 , 5; and ATP, 3 (standard conditions). Protein (myosin S1) concentration was 20 μg/ml. Incubation time was 5 min. Samples containing all components of the incubation medium without myosin S1 were taken as control of non-enzyme hydrolysis of ATP. The amount of inorganic phosphate released during ATP hydrolysis reaction was determined by the Chen method [26] by the measurement of optical absorbance of the solution at 820 nm using a microplate reader μQuwant (Biotek @ Instruments, Inc., USA) and specified as Pi nmol/min per 1 mg of protein.
The Zn 2+ and thiacalix[4]arene effects on the ATPase activity of myosin S1 were studied using standard incubation medium with solutions of ZnCl2 and thiacalix[4]arenes at the corresponding concentrations. The value of ATPase activity in the absence of ZnCl2 and/or calix[4]arenes in the incubation medium was taken as 100% (control).
The values of the imaginary constant of Michaelis (K m ) and maximal rate of myosin S1 ATPase for ATP (V max, ATP ) were calculated using the graph of the dependence of ATPase activity on the ATP concentration according to Lineweaver–Burk method [27]. Statistical processing of the obtained data was performed using standard methods of variation statistics. Experimental data were analyzed by using the standard software “MS Office” and “Statistica 4.5.” The statistical comparisons were performed using two-way analysis of variances (ANOVA).
Tetrahydroxy-thiacalix[4]arene-tetrasulphonate and tetrahydroxy-thiacalix[4]arene-tetraphosphonate were synthesized and characterized using NMR techniques and IR spectroscopy in the Phosphoranes Chemistry Department of the Institute of Organic Chemistry, NAS of Ukraine. Infrared and NMR spectroscopy confirmed the structure of these synthesized thiacalix[4]arenes. This thiacalix[4]arenes were dissolved in water.
Computer modeling of the interaction between ligands (thiacalix[4]arenes, Zn 2+ , model bindings) and receptor (myosin S1) was performed using AutoDock software, version 4.2 [28]. We used the three-dimensional enzyme structure with the 1b7t identifier in RSCB PDB in our research [29]. Computer modeling of the thiacalix[4]arene structural peculiarities was carried out using HyperChem 7.01. Molecular dynamics calculations were performed by the MM2 method with the semi-empirical methods (CNDO).
Program AutoDockTools was used for preliminary “processing” of interacting molecules. One hundred runs of Lamarkian genetic algorithms (population size, 100; the maximal number of energy evaluations, 10 6 ) were conducted. To analyze and visualize the docking results, we used the programs Chimera [30] and Yassara [31]. Calculation of the minimal total binding energy was implemented considering Van der Waals forces, electrostatic and hydrophobic interactions, and hydrogen bonds. The optimal ligand positions in the complex “receptor-ligand” were determined according to the energy values obtained by docking software calculator for binding energy in complex “receptor-ligand.” Thus, we selected a series of complexes with the lowest total energy and then calculated the optimal geometry of the complexes and determined the most energetically preferred arrangement of the ligands in the space of myosin subfragment-1 binding domain.
Tetrahydroxythiacalix[4]arene-tetrasulfosphonate
Tetrahydroxythiacalix[4]arene-tetraphosphonate
Complete Neglect of Differential Overlap (methods)
Michaelis constant, the substrate concentration at which the reaction rate of the enzyme is half of the maximal velocity
Lower 50-kDa domain of myosin
Lethal dose is the amount of an ingested substance that kills 50% of a test sample
A class of force fields
Myosin subfragment-1
National Academy of Science of Ukraine
Protein Data Bank
Phosphate-binding loop of myosin
Research Collaboratory for Structural Bioinformatics
The upper 50-kDa domain of myosin
Maximal velocity of the enzyme
V max for ATP
나노물질
초록 산화아연 나노입자(ZnO NPs)는 산업, 상업 제품 및 의약 분야를 포함한 광범위한 응용 분야에서 사용됩니다. ZnO NP의 독성에 대한 수많은 기계론적 연구가 깨끗한(신선한) NP에 대해 수행되었습니다. 그러나 변형된(노화) ZnO 나노입자에 의해 유도된 세포독성과 기본 메커니즘은 아직 명확하지 않습니다. 여기에서 우리는 시간이 지남에 따라 ZnO NP의 물리화학적 변형을 관찰한 후 신선하고 오래된 NP의 세포 독성을 평가했습니다. 우리는 신선한 ZnO 나노입자가 노화된 나노입자보다 더 높은 세포자멸사 수준을 유도한다는
3D 인쇄 프로젝트는 침대에서 나온 인쇄물에서 완벽하게 나올 수 있어야 하지만 대부분의 경우 동일한 재료가 아니기 때문에 그런 식으로 발생하지 않습니다. 완료하기 전에 프로젝트의 인쇄된 부분에 눈에 보이는 선과 불완전한 표면이 표시됩니다. 인쇄물이 보기 좋게 보이려면 마무리 공정을 적용해야 합니다. 거친 가장자리를 부드럽게 하고 인쇄물에서 확장된 필라멘트를 잘라 지지 재료를 제거한 후 습식 샌딩이 잘 작동합니다. 이것은 항상 지지 구조 또는 테두리의 결과이며 잘못 수행하면 인쇄물을 망칠 수 있습니다. 이미 완성된 3D 프린트