M1 대식세포 유래 엑소좀 MicroRNA-326은 매개 NF-κB 신호 전달 경로를 통해 간세포 암종 세포 진행을 억제합니다
초록
축적된 증거는 M1 대식세포 유래 엑소좀에서 유래한 마이크로RNA(miR)가 간세포 암종(HCC)의 진행을 조절할 수 있음을 보여주었습니다. 그러나 M1 대식세포 유래 엑소좀에서 유래한 miR-326이 간세포암종에 미치는 영향은 보고된 바 없다. 따라서, 본 연구의 목적은 HCC 세포 진행을 조절하는 M1 대식세포로부터 엑소좀 miR-326의 기전을 탐구하는 것이었다. RT-qPCR은 HCC 세포주에서 miR-326 발현을 감지했습니다. HCC에서 miR-326 발현은 형질감염에 의해 변경되었고, CD206 및 NF-κB 발현, 세포 증식, 집락 형성, 이동, 세포자멸사 및 침입에 대한 miR-326의 효과가 검출되었다. 이어서, 엑소좀을 M1 대식세포로부터 분리하였다. RT-qPCR은 M1 대식세포 유래 엑소좀에서 miR-326 발현을 확인했습니다. M1 대식세포 유래 엑소좀에서 miR-326 발현은 형질감염에 의해 변화되었다. M1 대식세포 유래 엑소좀을 HCC 세포와 공동 배양하여 HCC 세포의 생물학적 진행에 미치는 영향을 파악했습니다. 마지막으로 시험관 내 결과를 확인하기 위해 생체 내 실험을 수행했습니다. MiR-326은 HCC 세포에서 감소되었고 M1 대식세포 유래 엑소좀이 풍부했습니다. miR-326을 상향 조절하면 HCC 세포 증식, 집락 형성, 이동, 침입, CD206 및 NF-κB 발현을 억제하고 세포자멸사를 촉진하고 생체 내 HCC 종양의 성장을 억제합니다. , miR-326을 하향 조절하는 반면 반대 효과를 보였습니다. M1 대식세포 유래 엑소좀은 HCC 세포 증식, 콜로니 형성, 이동, 침입, CD206 및 NF-κB 발현을 억제하고 세포자멸사를 강화한 반면 miR-326의 과발현은 M1 대식세포 유래 엑소좀이 HCC 세포에 미치는 영향을 강화했습니다. M1 대식세포 유래 엑소좀 miR-326은 NF-κB 발현을 하향 조절함으로써 HCC의 증식, 이동 및 침입을 억제하고 세포자멸사를 진행시키는 것으로 밝혀졌습니다.
소개
간세포암종(HCC)은 전 세계적으로 다섯 번째로 흔한 암이며 가장 흔한 원발성 간암입니다[1]. 중국 국가 암 등록부(China National Cancer Registry)의 데이터에 따르면 원발성 간암의 사망률은 3위, 발병률은 4위입니다[2]. 간세포암종의 주요 위험인자는 C형 간염 바이러스와 B형 간염 바이러스에 의한 만성 감염, 아플라톡신에 오염된 식품, 비만, 흡연, 과음, 제2형 당뇨병이다[3]. 경동맥 화학색전술은 중기 간세포암종에 대한 확립된 치료법으로, 대부분의 중기 또는 진행기 간세포암종 환자의 생존율을 향상시킵니다[4]. 현재 간세포암종 진단은 주로 혈청 바이오마커와 영상기술에 의존한다[5]. 간세포암종 관련 5년 생존율은 약 60%에 불과하며 최근 몇 년 동안 발병률이 증가하고 있다[6]. 이를 감안할 때 간세포암종 치료에서는 정확한 치료 표적을 찾는 것이 최우선 과제입니다.
대식세포는 이펙터 세포이자 면역계의 주요 조절자이며 조직 리모델링 및 복구에 막대한 기능을 발휘하며 생체 내 거의 모든 조직에서 대사 기능을 조정합니다[7]. M1 대식세포는 종양 촉진 효과를 발휘하고 HCC 세포의 운동성을 향상시키는 것으로 밝혀졌습니다[8]. 엑소좀은 직경이 40~150nm인 원반 모양의 소포입니다[9]. Xu et al.에 따르면, 엑소좀 마이크로RNA(miRNA)는 표적 세포에서 유전자 발현을 조절함으로써 간세포암종의 증식, 침습성, 전이 및 약물 내성에 대한 기능을 갖는다[10]. 엑소좀을 함유한 miR-326이 다발성 경화증의 잠재적인 임상 표적이 될 수 있다는 연구 결과가 나왔다[11]. MiRNA는 특정 서열에서 mRNA 분해 및 번역 차단을 통해 많은 수의 단백질 암호화 유전자 발현을 조절함으로써 종양유전자 또는 종양 억제제로 작용할 수 있다[12]. 연구에서는 miR-326이 세포 주기 진행을 방해하고 세포자멸사를 강화하여 HCC 세포 성장을 억제하고, 상피-중간엽 전이 표현형을 감소시켜 세포 침습을 억제한다고 논의했습니다[13]. 또 다른 연구에서는 miR-326이 간세포암종 환자 치료를 위한 잠재적인 치료 표적이 될 수 있다고 보고했습니다[14]. 따라서 본 연구에서는 HCC 세포의 침입과 이동을 조절하는 엑소좀 miR-326의 메커니즘에 대해 논의했습니다.
자료 및 방법
윤리 성명서
모든 동물 실험은 국제 위원회의 실험 동물 관리 및 사용 지침을 준수했습니다. 프로토콜은 길림대학교 제3병원 기관 동물 관리 사용 위원회의 승인을 받았습니다.
대식세포 유도 및 식별
인간 단핵구 세포주 THP-1(Kunming Institute of Zoology, CAS, Kunming, China)은 10% 열-불활성화 소태아 혈청을 함유하는 RPMI 1640 배지(Gibco, CA, USA; Thermo Fisher Scientific, MA, USA)에서 배양되었습니다. FBS). THP-1 세포를 150ng/mL 포르볼 12-미리스테이트 13-아세테이트(PMA; P8139, Sigma-Aldrich, SF, CA, USA)와 반응시키고 RPMI 배지에서 24시간 동안 인큐베이션하여 M0 대식세포를 얻었다. 그런 다음 Wright 염색을 통해 유도 전과 후의 세포 형태를 관찰하였다. THP-1 세포 및 유도된 대식세포를 5 μL PBS에 재현탁하고 유리 슬라이드에 떨어뜨리고 Wright의 염료 용액으로 염색하고 완충 용액과 1:2로 혼합하였다. 10분 동안 염색하고 흐르는 물로 헹구고 현미경으로 세포를 관찰하였다. 또한, M0 대식세포 마커(CD68 및 CD206)는 역전사 정량적 중합효소 연쇄 반응(RT-qPCR)에 의해 측정되었습니다. 다음으로, 대식세포는 18시간 동안 20ng/mL Interferon(IFN)-γ(#285-IF; R&D Systems, MN, USA) 및 10pg/mL LPS(#8630; Sigma-Aldrich)와 함께 인큐베이션을 통해 M1 대식세포로 유도되었습니다. 시간. M1 대식세포 마커(IDO1 및 IL-12 p35)는 RT-qPCR로 검사했습니다[15].
엑소좀 추출
엑소좀 분리 키트(ExoEasy Maxi Kit, Qiagen, Hilden, Germany)로 엑소좀을 추출하였다. 대식세포 상층액을 무균 조건에서 15mL 원심분리관에 모아 0.8μM 여과막으로 여과하였다. 각 그룹의 세포 상층액에 XBP 완충액(1:1)을 첨가한 다음 500g에서 exoEasy 멤브레인 친화성 원심분리 컬럼으로 원심분리했습니다. . 세포에 10mL XWP 완충액을 보충하고 3000–5000g로 원심분리했습니다. . exoEasy 멤브레인 친화성 원심분리 컬럼을 400μL XE 용리 완충액으로 부화하고 500g에서 원심분리했습니다. . 용출 완충액을 exoEasy 멤브레인 친화성 원심분리 컬럼으로 옮기고 500g에서 원심분리했습니다. . 용리 완충액은 4 °C에서 24시간 동안 보관한 다음 식별에 사용했습니다. 나머지는 − 80 °C에서 보관되었습니다.
TEM 관찰 및 나노입자 추적 분석(NTA)
상기에서 얻은 엑소좀을 탄소 지지막 구리 메쉬에 떨어뜨리고 2% 인텅스텐산을 첨가하였다. 시료를 TEM으로 관찰하여 최적의 영상을 수집하여 분석하였습니다.
PBS의 불순물과 입자는 0.22μM 미세다공성 필터로 제거하였다. 엑소좀의 입자 밀도에 따라 여과된 PBS를 적절한 농도로 희석하고 Nanosight NS300 나노입자 검출기(Malvern, Westborough, MA, USA)를 사용하여 검출하였다.
식별 후, miR-326 억제제 및 miR-326 억제제 음성 대조군(NC), miR-326 모방체 및 miR-326 모방체 NC로 형질감염된 대식세포 유래 엑소좀을 엑소좀 분리 키트(Invitrogen)로 추출했습니다.
RT-qPCR
총 RNA는 Trizol Reagent(Thermo Fisher)로 추출하였고, Real-Time PCR은 SYBR-Green PCR Master Mix(Roche)와 ABI 7500 Real-Time PCR System(Life Technologies, Grand Island, NY, USA)을 이용하여 수행하였다. ). 프라이머 염기서열은 Table 1과 같다. 정량분석은 2
-△△Ct
의 방법으로 진행하였다. .
서부 얼룩 분석
세포 및 엑소좀의 총 단백질을 추출하였다. 단백질 농도는 BCA(bicinchoninic acid) 키트(Boster Biological Technology Co. Ltd., 무한, 후베이, 중국)에 의해 결정되었습니다. 단백질에 샘플 버퍼를 첨가하고 95 °C에서 끓이고 각 웰에 30 μg을 로딩했습니다. 단백질을 10% 폴리아크릴아미드 겔 전기영동(Boster Biological Technology)으로 분리하고 폴리비닐리덴 플루오라이드 멤브레인에 전기 블로팅한 후 5% 소 혈청 알부민(BSA)으로 밀봉하였다. 막은 CD63(1:1000, Developmental Studies Hybridoma Bank, University of Iowa, Ames, IA, USA), CD181(1:1000, R&D Systems), GAPDH(1:2000, Jackson ImmunoResearch Laboratories, PA, USA) 및 양고추냉이 과산화효소로 표지된 이차 항체(1:500, Jackson ImmunoResearch Laboratories)로. 이미지는 Odyssey 듀얼 컬러 적외선 형광 스캐닝 이미징 시스템으로 얻었고 밴드의 회색 값은 Quantity One 이미지 분석 소프트웨어로 측정했습니다.
세포 배양 및 스크리닝
정상 간 세포주 HL-7702 및 인간 HCC 세포주 BEL-7404, HepG2, SMMC-7721 및 QGY-7703을 선택하고 10% 소태아혈청(FBS), 페니실린(100U/mL)이 포함된 Gibco RPMI Media 1640에서 배양했습니다. ) 및 스트렙토마이신(100mg/mL). MiR-326 발현은 RT-qPCR에 의해 검출되었으며, 적합한 세포주를 스크리닝하였다.
엑소좀 라벨링 및 엑소좀 흡수
엑소좀은 250μL 희석제 C로 재현탁되었고 엑소좀 막이 손상되지 않도록 부드럽게 분쇄되었습니다. PKH67 염료(1μL, Sigma-Aldrich)를 250μL Diluent C에 첨가하여 500μL에 도달하도록 하고 인큐베이션했습니다. 용액에 500μL 1% BSA를 첨가하고 37°C에서 1분 동안 인큐베이션했습니다. 120,000g에서 원심분리하여 엑소좀을 얻었다. 2시간 동안 4°C PKH67로 표지된 엑소좀은 120,000g에서 원심분리하여 얻었습니다. 2시간 동안 4°C 엑소좀은 빛을 피하는 6mL RPMI-1640 배지로 재현탁되었습니다. 그런 다음, 표지된 엑소좀을 12시간 동안 HCC 세포와 공동 배양하였다. 이후 배양액을 제거하고 PBS로 5분/회 3회 세척하고 간세포암종 세포에 내부적으로 흡수되지 않은 형광 표지 엑소좀을 충분히 씻어내었다. 엑소좀은 4% 파라포름알데히드로 고정하고 4'-6-디아미디노-2-페닐인돌로 염색하였다. 밀봉 후 레이저 공초점 현미경으로 형광 분포를 관찰하였다.
셀 그룹화 및 처리
HepG2 세포와 SMMC-7721 세포를 12웰 플레이트에 0.5–1 × 10
6
으로 시딩했습니다. 세포/웰. 50-60% 합류로 세포를 Lipofectamine 2000(Invitrogen, Carlsbad, CA)으로 형질감염시켰다. HepG2 세포는 miR-326-모방 그룹(miR-326 모방으로 형질감염됨) 및 NC-모방 그룹(miR-326 모방 NC로 형질감염됨)으로 분포되었다. SMMC-7721 세포는 miR-326 억제제 그룹(miR-326 억제제로 형질전환됨)과 NC 억제제 그룹(miR-326 억제제 NC로 형질전환됨)으로 할당되었습니다. miR-326-모방체, miR-326-억제제 및 이들의 NC는 형질감염을 위해 Lipofectamine 2000과 혼합되었습니다. 처리하지 않은 HepG2 세포와 SMMC-7721 세포를 공백군으로 설정하였다. miR-326-mimic, miR-326-inhibitor 및 그들의 NC는 Guangzhou RibBio Co., Ltd.(Guangzhou, China)에서 고안하고 구성했습니다(표 1).
간세포암종 세포와 M1 대식세포 유래 엑소좀의 공동 배양
M1 대식세포 유래 엑소좀 현탁액의 단백질 농도를 BCA 방법으로 검출하고, 50 μg 단백질을 포함하는 상응하는 엑소좀 현탁액의 부피를 계산하였다. HepG2 세포 및 SMMC-7721 세포를 1 × 10
5
의 12웰 플레이트에 접종했습니다. 웰당 세포/mL. HepG2 세포는 대조군(Exosome과 함께 배양되지 않은 HepG2 세포), 엑소좀(Exo) 그룹(M1 대식세포 유래 엑소좀과 공동 배양된 HepG2 세포), Exo-miR-326-모방체(HepG2 miR-326 모방체로 형질감염된 M1 대식세포 유래 엑소좀과 공동 배양된 세포), 엑소-NC-모방 그룹(miR-326 모방체로 형질감염된 M1 대식세포 유래 엑소좀과 공동 배양된 HepG2 세포). SMMC-7721 세포는 또한 4개의 그룹으로 할당되었다:블랭크 그룹(SMMC-7721 세포는 엑소좀과 공동 배양되지 않음), 엑소 그룹(SMMC-7721 세포는 M1 대식세포 유래 엑소좀과 공동 배양됨), Exo-miR-326- Inhibitor group (SMMC-7721 cells with co-cultured with M1 macrophage-derived exosome that transfected with miR-326 inhibitor), Exo-NC-inhibitor group (SMMC-7721 cells with miR-transfected M1 macrophage-derived exosomes co-cultured) 326 억제제 NC).
3-(4,5-디메틸티아졸-2-일)-2,5-디페닐테트라졸륨 브로마이드(MTT) 분석
세포를 트립신으로 분리하고 세포 밀도가 4 × 10
4
인 96웰 플레이트에 접종했습니다. 웰당 세포. 배양 배지는 각각 12, 24, 36, 48, 60시간 배양 후 폐기하였다. 500μL 0.5g/L MTT 용액과 함께 인큐베이션한 후 세포에 200μL 디메틸 설폭사이드 용액을 첨가하고 분쇄하고 부화했습니다. 광학 밀도(OD, 490 nm) 값은 마이크로플레이트 리더로 측정했습니다.
콜로니 형성 분석
24시간 동안 배양하고 트립신으로 분리한 세포를 접시당 300개의 세포가 있는 35mm 작은 접시에 접종했습니다. 용액은 3일마다 교체되었습니다. 10일 배양 후 세포를 40g/L
-1
로 고정 파라포름알데히드 및 1g/L
−1
로 염색 크리스탈 바이올렛 용액 및 건조. 콜로니 수(50개 이상의 세포)를 현미경으로 계산했습니다.
트랜스웰 분석
셀(1 × 10
5
) )를 200μL 블랭크 배양 배지로 현탁시켰다. 실험은 Transwell chamber(Corning Glass Works, Corning, N.Y., USA)의 지침에 따라 수행되었습니다(침습 실험에는 matrigel이 필요했지만 마이그레이션 실험에는 필요하지 않음). RPIM 1640(10% FBS, 600μL)을 하부 챔버에 추가했습니다. 상부 및 하부 챔버는 matrigel(BD Biosciences, Franklin Lakes, NJ, USA)로 미리 코팅된 Transwell 멤브레인으로 분리되었습니다. 24시간 동안 배양한 후, 챔버를 95% 알코올로 고정하였다. 크리스탈 바이올렛 용액으로 염색한 후 현미경으로 5개의 시야에서 세포를 관찰했습니다.
유세포분석
세포 주기:트립신에 의해 세포가 분리되었습니다. 셀(1 × 10
6
) )을 0.5 mL PBS로 현탁시키고 단일 현탁액으로 분쇄하였다. 얼음 위에서 미리 냉각된 70% 에탄올 4.5mL와 혼합하고, 세포를 3000g에서 원심분리했습니다. , 5mL PBS로 헹구고 3000g에서 다시 원심분리 . 이어서, 세포를 0.2 mg RnaseA를 함유하는 1 mL PI/Triton X-100 염색 용액(20 μg PI/0.1% Triton X-100)으로 현탁시켰다. 세포 주기는 유세포 분석으로 감지되었습니다.
세포 사멸:트립신 처리된 세포(1 × 10
6
)을 1mL PBS로 현탁시키고, 3000g에서 분쇄하고 원심분리했습니다. . 세포를 배양 완충액(10mmol/L Hepes/NaOH, pH 7.4, 140mmol/L NaCl, 5mmol/L CaCl2)으로 차례로 헹구었습니다. ) 및 3000g에서 원심분리 . 다음으로, 세포를 100μL 마킹 용액(FITC-Annexin V 및 PI가 1μg/mL에 도달하도록 인큐베이션 버퍼에 첨가)과 함께 인큐베이션하고 3000g에서 원심분리했습니다. , 배양 완충액으로 한 번 세척하고 형광성(SA-FLOUS) 용액으로 부화합니다. 세포 사멸은 유세포 분석에 의해 감지되었습니다. 유세포분석기의 파장은 488 nm였으며, FITC 형광은 515 nm에서 밴드 필터에 의해 검출되었고, PI는 560 nm보다 큰 파장에서 검출되었다. 결과는 컴퓨터에 의해 자동으로 분석되었습니다.
누드 마우스의 종양 이종이식
4~6세의 생쥐 40마리(중국 길림대학교 동물과학대학)를 각 그룹에 5마리씩 8개 그룹으로 무작위로 분배했습니다. 마우스는 특정 병원균이 없는 등급의 동물 실험실에서 1w 동안 먹였으며, 사료, 쿠션 및 물병은 제때에 교체되었습니다. 생쥐의 건강 상태는 매일 관찰되어야 합니다. 1주일 후, HCC 세포를 세포 현탁액으로 제조하고 0.1mL 세포 현탁액(1 × 10
6
)으로 목과 등에 피하 주사했습니다. ). 종양의 성장은 3-5일 후에 관찰되었습니다. 누드 마우스의 무게를 4일마다 측정하고 종양 부피를 측정했습니다. 누드 마우스는 주사 20일 후에 안락사되었습니다.
통계 분석
모든 데이터는 SPSS 17.0 소프트웨어(SPSS Statistics, Chicago, IL, USA)로 해석되었습니다. 측정 데이터는 평균 ± 표준편차로 표시하였다. 두 그룹 간의 비교는 t -테스트, 여러 그룹 간의 비교는 일원 분산 분석(ANOVA)에 의해 평가되었습니다. 피 value <0.05는 통계적으로 유의한 차이를 나타냅니다.
<섹션 데이터-제목="결과">
결과
M1 대식세포 및 엑소좀의 식별
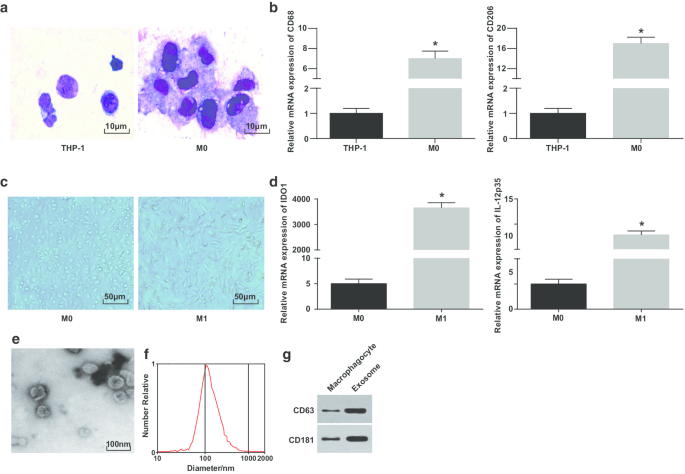
Wright 염색을 이용하여 PMA에 의해 유도된 THP-1 세포의 형태를 관찰하였다. 유도 전의 THP-1 세포의 부피는 적었고 caryoplasm의 비율은 더 높았습니다. 유도 후 세포의 형태는 불규칙하고 부피는 더 커지고 핵질의 비율은 감소했습니다. 세포질은 입자가 풍부하고 액포가 적은 밝은 파란색을 띠고 더 풍부했습니다. 핵은 자주색을 띠고 종종 한쪽으로 기울어져 있어 세포가 대식세포의 전형적인 형태적 특징을 가지고 있음을 보여줍니다(그림 1a).
<그림>
M1 대식세포 및 엑소좀의 식별. 아 PMA 처리 전후에 THP-1 세포의 형태를 관찰하기 위한 Wright 염색. ㄴ RT-qPCR에 의해 검출된 THP-1 세포 및 PMA 처리된 THP-1 세포에서의 CD68 및 CD206 발현; ㄷ 대식세포와 M1 대식세포의 형태. d RT-qPCR에 의해 검출된 M0 대식세포 및 LPS 및 INF-γ 처리된 M0 대식세포에서 IDO1 및 IL-12 p35의 발현. 이 엑소좀 관찰을 위한 TEM 에 NTA에 의한 엑소좀의 입자 크기 분포 검출. 지 CD63 및 CD181의 단백질 밴드. 패널 b에서 *P <0.05 대 THP-1 세포; 패널 d에서 *P <0.05 대 M0 대식세포. 측정 데이터는 평균 ± 표준편차(N =3), 두 그룹 간의 비교는 t 검정에 의해 수행되었습니다.
대식세포의 성공적인 유도를 추가로 확인하기 위해 유도 전후의 CD68 및 CD206 발현을 RT-qPCR로 테스트했습니다. PMA 유도 이후 CD68 및 CD206 발현이 상승한 것으로 나타났으며, 이는 PMA가 THP-1 세포를 M0 대식세포로 성공적으로 유도했음을 나타냅니다(그림 1b). 그런 다음, M0 대식세포는 LPS 및 INF-γ의 유도에 의해 M1 대식세포로 분극화되었다. 대식세포의 형태, M1형 대식세포 표면 마커 IDO1 및 IL-12 p35 발현을 관찰하고 시험하였다. M0 대식세포는 원형, 타원형 또는 방추형으로 다양하고 불규칙한 부착 형태를 보였다. IFN-γ에 의해 자극된 대식세포의 형태는 더 많은 pseudopods와 돌출부 및 fusiformis로 입증되었습니다(그림 1c). RT-qPCR은 LPS 및 INF-γ 처리 후 M1 대식세포가 마커(IDO1 및 IL-12p35)의 증가를 나타냈다고 설명했습니다(그림 1d).
이어서 대식세포에서 유래한 엑소좀을 TEM으로 관찰하였다. 대식세포에서 유래한 엑소좀은 풍부하고 모양이 원형 또는 타원형이며 막 구조, 균일한 크기 및 오염 물질이 적은 것으로 나타났습니다(그림 1e). NTA는 MODE 곡선의 중앙 집중식 피크와 부드러운 선형이 있는 엑소좀이 더 집중된 직경과 더 적은 불순물을 가짐을 표시했습니다(그림 1f). 웨스턴 블롯 분석은 대식세포와 비교하여 특정 마커 단백질 CD63 및 CD181 발현이 대식세포에서 유래된 엑소좀에서 증가했다고 보고했습니다(그림 1g). 이러한 결과는 단핵구가 대식세포로 분화되도록 성공적으로 유도하여 M1 대식세포로 분극화되었음을 나타냅니다.
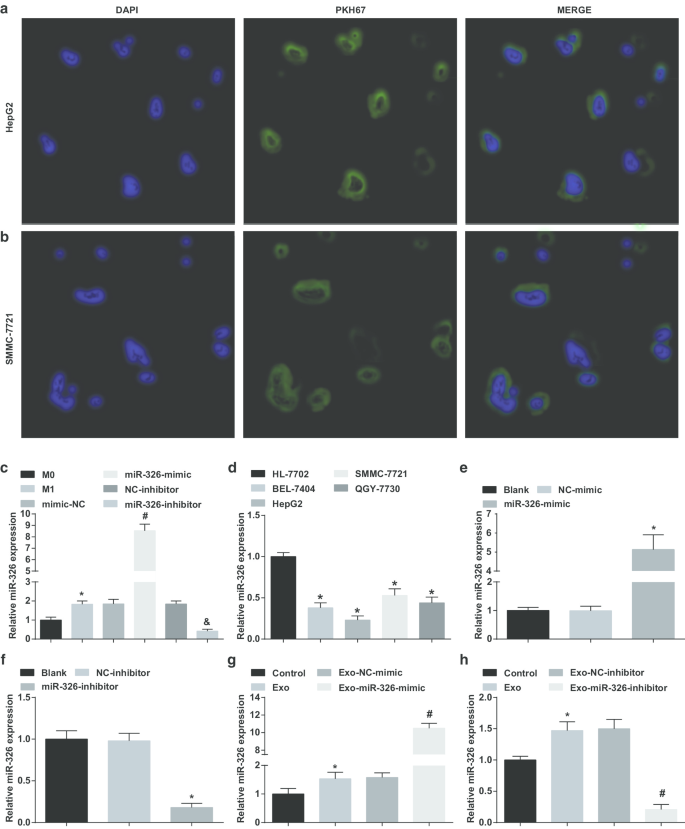
M1 대식세포에서 유래된 엑소좀이 miR-326을 HCC 세포로 수송했는지 확인하기 위해 HepG2 및 SMMC-7721 세포를 엑소좀과 공동 배양하였다. 형광현미경하에서 4시간 transfection했을 때 많은 수의 엑소좀이 HepG2와 SMMC-7721 세포에 동화되었음을 알 수 있었다(Fig. 2a, b).
<그림>
M1 대식세포 유래 엑소좀은 miR-326을 HCC 세포에 전달하고 HCC 세포에서 miR-326 발현에 영향을 미칩니다. 아 HepG2 세포에 의한 M1 대식세포 유래 엑소좀의 흡수. ㄴ SMMC-7721 세포에 의한 M1 대식세포 유래 엑소좀의 흡수. ㄷ INF-γ 및 LPS 유도 전과 후의 대식세포 엑소좀의 miR-326 발현 비교. d RT-qPCR에 의해 검출된 HCC 세포주(BEL-7404, HepG2, SMMC-7721, QGY-7703) 및 인간 정상 간세포 HL-7702 세포주에서의 miR-326 발현. 이 RT-qPCR은 HepG2 세포에서 miR-326 발현에 대한 miR-326 모방의 효과를 감지했습니다. 에 RT-qPCR은 SMMC-7721 세포에서 miR-326 발현에 대한 miR-326 억제제의 효과를 감지했습니다. 지 RT-qPCR은 MiR-326 모방 형질감염 M1 대식세포 유래 엑소좀이 HepG2 세포에서 miR-326 발현에 미치는 영향을 감지했습니다. 아 RT-qPCR은 SMMC-7721 세포에서 miR-326 발현에 대한 miR-326 억제제 형질감염 M1 대식세포 유래 엑소좀의 효과를 검출했습니다. 패널 c에서 *P <0.05 대 M0 대식세포; 패널 d에서 *P <0.05 대 HL-7702 세포; 패널 e에서 *P <0.05 대 NC 모방 그룹; 패널 f에서 *P <0.05 대 NC-억제제 그룹; 패널 g에서 *P <0.05 대 대조군, #P <0.05 대 엑소-NC-모방 그룹; 패널 h에서 *P <0.05 대 대조군, #P <0.05 대 Exo-NC-억제제 그룹. 측정 데이터는 평균 ± 표준편차(N =3), 다중 그룹 간의 비교는 분산의 일원 분석에 의해 수행되었습니다.
그런 다음 HCC 세포주에서 miR-326 발현을 테스트했습니다. 나타난 바와 같이, miR-326 발현은 HL-7702 세포에 비해 BEL-7404, HepG2, SMMC-7721 및 QGY-7703 세포에서 감소되었으며, 그 중 HepG2 세포가 가장 낮은 발현을 나타내었고 SMMC-7721 세포에서 가장 높은 발현을 나타냈다. (그림 2d).
이어서 miR-326-mimic 및 miR-326-inhibitor를 HepG2 및 SSMC-7721 세포에 각각 형질감염시켜 miR-326 발현에 미치는 영향을 조사하였다. MiR-326은 HepG2 세포에서 증가된 miR-326 발현을 모방한 반면, miR-326 억제제는 SSMC-7721 세포에서 miR-326 발현을 감소시켰습니다(그림 2e, f).
다음으로, miR-326-모방체 및 miR-326-억제제로 형질감염된 M1 대식세포의 엑소좀을 각각 HepG2 및 SSMC-7721 세포와 공동 배양하였다. 엑소좀과의 공동 배양은 HCC 세포에서 miR-326 발현을 증가시켰고, miR-326-모방 형질감염된 M1 대식세포 유래 엑소좀은 HepG2 세포에서 miR-326 발현을 추가로 증가시킨 반면, miR-326-억제제-형질감염된 M1 대식세포는 강조되었다 -유래 엑소좀은 SSMC-7721 세포에서 miR-326 발현을 저하시켰다(그림 2g, h). M1 대식세포 유래 엑소좀은 miR-326을 HCC 세포에 전달하고 HCC 세포에서 miR-326 발현에 영향을 미치는 것으로 제안됩니다.
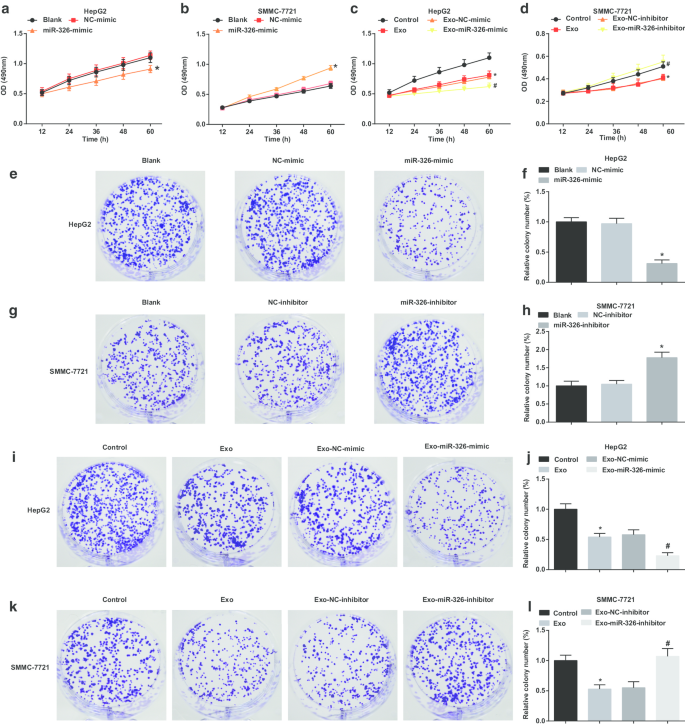
M1 대식세포 유래 엑소좀 miR-326은 HCC 세포에서 세포 증식 및 집락 형성 능력을 감소시킵니다.
HCC 세포 증식에 대한 엑소좀 miR-326의 영향을 조사하기 위해 MTT 및 콜로니 형성 분석을 수행하여 HCC 세포 증식을 조사하였다. HepG2 세포에서 miR-326 복원은 세포 증식과 집락 형성 능력을 손상시키는 것으로 제안되었습니다(그림 3a, e, f). M1 대식세포 유래 엑소좀은 HepG2 세포가 증식하고 식민지를 형성하는 것을 방해했습니다. MiR-326 모방 형질감염 M1 대식세포 유래 엑소좀은 세포 증식 및 집락 형성 능력을 더욱 손상시킵니다(그림 3c, i, j).
<그림>
M1 대식세포 엑소좀 miR-326은 HCC 세포에서 세포 증식과 집락 형성을 억제합니다. 아 MTT 분석은 MiR-326 모방체의 형질감염이 HepG2 세포의 증식에 미치는 영향을 감지했습니다. ㄴ MTT 분석은 SMMC-7721 세포의 증식에 대한 miR-326 억제제의 형질감염 효과를 검출하였다. ㄷ MTT 분석은 MiR-326 모방 형질감염 M1 대식세포 유래 엑소좀과의 공동 배양이 HepG2 세포의 증식에 미치는 영향을 감지했습니다. d MTT 분석은 SMMC-7721 세포의 증식에 대한 miR-326 억제제로 형질감염된 M1 대식세포 유래 엑소좀과의 공동 배양 효과를 감지했습니다. 이 집락 형성 분석은 MiR-326 모방체의 형질감염이 HepG2 세포의 집락 형성 능력에 미치는 영향을 검출했습니다. 에 HepG2 세포의 콜로니 수. 지 집락 형성 분석은 SMMC-7721 세포의 집락 형성 능력에 대한 miR-326 억제제의 형질감염 효과를 검출하였다. 아 SMMC-7721 세포의 콜로니 수. 나 집락 형성 분석은 HepG2 세포의 집락 형성 능력에 대한 miR-326 모방 형질감염 M1 대식세포 유래 엑소좀과의 공동 배양 효과를 검출했습니다. j 엑소좀으로 처리된 HepG2 세포의 콜로니 수. 케이 집락 형성 분석은 SMMC-7721 세포의 집락 형성 능력에 대한 miR-326 억제제 형질감염 M1 대식세포 유래 엑소좀의 공동 배양 효과를 검출했다. 나 엑소좀으로 처리된 SMMC-7721 세포의 콜로니 수. 패널 및 f에서 *P <0.05 대 NC 모방 그룹; 패널 b와 h에서 *P <0.05 대 NC-억제제 그룹; 패널 c와 j에서 *P <0.05 대 대조군, #P <0.05 대 엑소-NC-모방 그룹; 패널 d와 l에서 *P <0.05 대 대조군, #P <0.05 대 Exo-NC-억제제 그룹. 측정 데이터는 평균 ± 표준편차(N =3), 다중 그룹 간의 비교는 분산의 일원 분석에 의해 수행되었습니다.
SMMC-7721 세포에서 miR-326 녹다운은 세포 증식과 집락 형성 능력을 향상시켰다(그림 3b, g, h). M1 대식세포 유래 엑소좀을 처리한 SMMC-7721 세포에서는 세포 증식과 집락 형성 능력이 감소하였다. MiR-326-억제제-형질전환된 M1 대식세포 유래 엑소좀은 세포 증식 및 집락 형성 능력을 더욱 촉진시켰다(그림 3d, k, l). M1 대식세포 유래 엑소좀에서 유래한 miR-326은 간세포암종 세포의 증식을 방해한다는 암시가 있습니다.
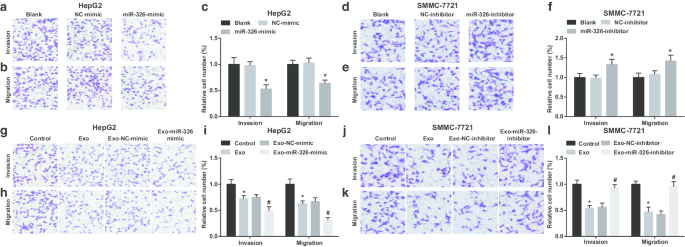
M1 대식세포 유래 엑소좀 miR-326은 HCC 세포의 이동 및 침범을 억제합니다.
그런 다음, 엑소좀 miR-326이 간세포암종 세포의 침습 및 이동에 미치는 영향을 조사하였다. HepG2 세포에서 miR-326의 복원이 침입과 이동을 제한한다는 것이 입증되었습니다(그림 4a-c). M1 대식세포 유래 엑소좀은 HepG2 세포가 침입하여 이동하도록 방해했습니다. 침습 및 이동은 HepG2 세포가 miR-326-모방 형질감염된 M1 대식세포 유래 엑소좀과 공동 배양될 때 더욱 저하되었습니다(그림 4g-i).
<그림>
M1 대식세포 엑소좀 miR-326은 HCC 세포의 이동과 침입을 억제합니다. 아 Transwell 분석은 HepG2 세포의 침입에 대한 miR-326 모방체의 형질감염 효과를 감지했습니다. ㄴ Transwell 분석은 HepG2 세포의 이동을 모방한 miR-326의 형질감염 효과를 감지했습니다. ㄷ HepG2 세포의 침입 및 이동 수. d Transwell 분석은 SMMC-7721 세포의 침입에 대한 miR-326 억제제의 형질 감염 효과를 감지했습니다. 이 Transwell 분석은 SMMC-7721 세포의 이동에 대한 miR-326 억제제의 형질감염 효과를 감지했습니다. 에 SMMC-7721 세포의 침입 및 이동 수. 지 Transwell 분석은 MiR-326 모방 형질감염 M1 대식세포 유래 엑소좀과의 공동 배양이 HepG2 세포의 침입에 미치는 영향을 감지했습니다. 아 Transwell 분석은 MiR-326 모방 형질감염 M1 대식세포 유래 엑소좀과의 공동 배양이 HepG2 세포의 이동에 미치는 영향을 감지했습니다. 나 M1 대식세포 유래 엑소좀과 공배양된 HepG2 세포의 침입 및 이동 수. j Transwell 분석은 SMMC-7721 세포의 침입에 대한 miR-326 억제제 형질감염 M1 대식세포 유래 엑소좀과의 공동 배양 효과를 감지했습니다. 케이 Transwell 분석은 SMMC-7721 세포의 이동에 대한 miR-326 억제제 형질감염 M1 대식세포 유래 엑소좀과의 공동 배양 효과를 감지했습니다. 나 M1 대식세포 유래 엑소좀과 공배양된 SMMC-7721 세포의 침입 및 이동 수. 패널 c에서 *P <0.05 대 NC 모방 그룹; 패널 f에서 *P <0.05 대 NC-억제제 그룹; 패널 i에서 *P <0.05 대 대조군, #P <0.05 대 엑소-NC-모방 그룹; 패널 l에서 *P <0.05 대 대조군, #P <0.05 대 엑소-NC-억제제 그룹. Measurement data were depicted as mean ± standard deviation (N = 3), comparisons among multiple groups were conducted by one-way analysis of variance
MiR-326 knockdown resulted in enhancements in SMMC-7721 cell invasion and migration (Fig. 4d–f). When treated with M1 macrophages-derived exosomes, SMMC-7721 cells were exhibited with decreased invasion and migration. However, SMMC-7721 cell invasion and migration were boosted upon co-culture with miR-326-inhibitor-transfected M1 macrophages-derived exosomes (Fig. 4j–l). It is implied that miR-326 derived from M1 macrophage exosomes impedes the invasion and migration of HCC cells.
M1 Macrophage-Derived Exosomal miR-326 Promotes Apoptosis of HCC Cells
When examining the effect of exosomal miR-326 on the cell cycle and apoptosis of HCC cells, PI single staining and Annexin V-FITC/PI double staining were applied. It was illustrated that miR-326 overexpression increased cells arrested at G0/G1 phase, reduced cells arrested at S and G2/M phases and raised apoptosis in HepG2 cells (Fig. 5a–d). Co-culturing with M1 macrophages-derived exosomes increased cells arrested at G0/G1 phase, reduced cells arrested at S and G2/M phases and raised cell apoptosis of HepG2 cells. Co-cultivation with exosomes from M1 macrophages transfected with miR-326-mimic further enhanced these effects (Fig. 5i–l).
M1 macrophage exosomal miR-326 promotes apoptosis of HCC cells. 아 , b Flow cytometry detected the effect of transfection of miR-326 mimic on HepG2 cell cycle; ㄷ , d flow cytometry detected the effect of transfection of miR-326 mimic on HepG2 cell apoptosis; 이 , f flow cytometry detected the effect of transfection of miR-326 inhibitor on SMMC-7721 cell cycle; 지 , h flow cytometry detected the effect of transfection of miR-326 inhibitor on SMMC-7721 cell apoptosis; i , j flow cytometry detected the effects of co-culture of with miR-326 mimic-transfected M1 macrophage-derived exosomes on HepG2 cell cycle; 케이 , 나 flow cytometry detected the effect of co-culture with miR-326 mimic-transfected M1 macrophage-derived exosomes on HepG2 cell apoptosis; m , n flow cytometry detected the effect of co-culture with miR-326 inhibitor-transfected M1 macrophage-derived exosomes on SMMC-7721 cell cycle; o , p , flow cytometry detected the effect of co-culture with miR-326 inhibitor-transfected M1 macrophage-derived exosomes on SMMC-7721 cell apoptosis. In panel b and d, *P <0.05 versus the NC-mimic group; In panel f and h, *P <0.05 versus the NC-inhibitor group; In panel j and l, *P <0.05 versus the control group, #P <0.05 versus the Exo-NC-mimic group; In panel n and p, *P <0.05 versus the controlgroup, #P <0.05 versus the Exo-NC-inhibitor group. Measurement data were depicted as mean ± standard deviation (N = 3), comparisons among multiple groups were conducted by one-way analysis of variance
In SMMC-7721 cells, miR-326 down-regulation reduced cells arrested at G0/G1 phase, elevated cells arrested at S and G2/M phases, and declined cell apoptosis (Fig. 5e–h). Untransfected M1 macrophages-derived exosomes increased cells arrested at G0/G1 phase, reduced cells arrested at S and G2/M phases, and heightened cell apoptosis. MiR-326-inhibitor-transfected M1 macrophages-derived exosomes degraded cells arrested at G0/G1 phase, elevated cells arrested at S and G2/M phases, and decreased cell apoptosis (Fig. 5m–p). Briefly, it is summarized that miR-326 derived from M1 macrophage exosomes arrests cell cycle in G0/G1 phase and induces cell apoptosis in HCC.
M1 Macrophage-Derived Exosomal miR-326 Declines CD206 and NF-κB Expression in HCC Cells
Next, the potential mechanism of miR-326 derived from M1 macrophage exosomes in the biological progress of HCC cells was explored. NF-κB is the key link between inflammation and cancer. Many regulatory proteins and miRNAs could inhibit the excessively activated NF-κB signaling to suppress cancer [16]. Such a beneficial effect may include the polarization of M2 macrophages into M1 macrophages. CD206 and NF-κB expression in HepG2 and SMMC-7721 cells was tested by RT-qPCR. It was suggested that miR-326 restoration decreased CD206 and NF-κB expression in HepG2 cells, while miR-326 knockdown enhanced CD206 and NF-κB expression in SMMC-7721 cells (Fig. 6a, b). Moreover, co-culture with M1 macrophage exosomes significantly reduced CD206 and NF-κB expression in HepG2 cells, while co-culture with M1 macrophage exosomes-overexpressing miR-326 further decreased CD206 and NF-κB expression. Treated with untransfected M1 macrophages-derived exosomes, CD206 and NF-κB expression was decreased in SMMC-7721 cells. Co-cultured with miR-326-inhibitor-transfected M1 macrophages-derived exosomes, SMMC-7721 cells were featured by heightened CD206 and NF-κB expression (Fig. 6c, d). It was concluded that miR-326 from M1 macrophage exosomes played a tumor suppressor by inhibiting NF-κB in HCC cells.
M1 macrophage exosomal miR-326 declines CD206 and NF-κB expression in HCC cells. 아 RT-qPCR detected the effect of transfection of miR-326 mimic on the expression of CD206 and NF-κB in HepG2 cells; ㄴ RT-qPCR detected the effect of transfection of miR-326 inhibitor on the expression of CD206 and NF-κB in SMMC-7721 cells. ㄷ RT-qPCR detected the effect of co-culture with miR-326 mimic-transfected M1 macrophage-derived exosomes on the expression of CD206 and NF-κB in HepG2 cells; d RT-qPCR detected the effect of co-culture with miR-326 inhibitor-transfected M1 macrophage-derived exosomes on the expression of CD206 and NF-κB in SMMC-7721 cells. In panel a, *P <0.05 versus the NC-mimic group; In panel b, *P <0.05 versus the NC-inhibitor group; In panel c, *P <0.05 versus the control group, #P <0.05 versus the Exo-NC-mimic group; In panel d, *P <0.05 versus the control group, #P <0.05 versus the Exo-NC-inhibitor group. Measurement data were depicted as mean ± standard deviation (N = 3), comparisons among multiple groups were conducted by one-way analysis of variance
miR-326 from M1 Macrophage Exosomes Inhibits HCC Tumor Growth In Vivo
Finally, the in vivo results were validated through tumor xenografts. As displayed, miR-326 overexpression decreased volume and weight of tumors in HepG2 cells (Fig. 7a–c). In mice transplanted with HepG2 cells co-cultured with exosomes, the treatment with M1 macrophage exosomes significantly reduced the tumor volume and weight of HepG2 cells, while treatment with M1 macrophages-overexpressing miR-326 further reduced the tumor volume and weight of HepG2 cells (Fig. 7g–i).
M1 macrophage exosomal miR-326 reduces the volume and weight of HCC tumor in vivo. 아 –ㄷ The effect of transfection of miR-326 mimic on the tumor volume and tumor of nude mice xenografted with HepG2 cells. d –f The effect of transfection of miR-326 inhibitor on the tumor volume and tumors of nude mice xenografted with SMMC-7721 cells. 지 –나 The effect of co-culture with miR-326 mimic-transfected M1 macrophage-derived exosomes on the tumor volume and tumor of nude mice xenografted with HepG2 cells. j –l The effect of co-culture with miR-326 inhibitor-transfected M1 macrophage-derived exosomes on the tumor volume and tumor of nude mice xenografted with SMMC-7721 cells. In panel b and c, *P <0.05 versus the NC-mimic group; In panel e and f, *P <0.05 versus the NC-inhibitor group; In panel h and i, *P <0.05 versus the control group, #P <0.05 versus the Exo-NC-mimic group; In panel k and l, *P <0.05 versus the control group, #P <0.05 versus the Exo-NC-inhibitor group. Measurement data were depicted as mean ± standard deviation (n = 5), comparisons among multiple groups were conducted by one-way analysis of variance
In SMMC-7721 cells, miR-326 suppression increased volume and weight of tumors (Fig. 7d–f). Untransfected M1 macrophages-derived exosomes obstructed tumor growth in volume and weight of SMMC-7721 cells. Co-cultured with miR-326 inhibitor-transfected M1 macrophages-derived exosomes, SMMC-7721 cells were injected into mice and caused elevations in tumor volume and weight (Fig. 7j–l). Summarily, miR-326 derived from M1 macrophage exosomes depressed tumor growth of HCC in vivo.
Discussion
HCC is a common cancer that is characterized with high morbidity and mortality, difficult early diagnosis and treatment, poor prognosis and 5-year survival rate [17]. Recently, a study has highlighted that the lowly expressed lncRNA cox-2 declines the ability of M1 macrophages to suppress HCC cell growth, invasiveness, angiogenesis migration and promote apoptosis [18]. Hu et al. have discussed that miR-326 is obviously degraded in HCC tissues and cell lines, while down-regulated miR-326 is connected to the TNM stage, lymph node metastasis and differentiation of HCC patients [14]. It is customarily considered that HCC cells-derived exosomes can form a fertile environment to facilitate HCC cells growth, invasiveness and metastasis as well as development of drug resistance [19]. The current study was designed to explore the mechanism of exosomal miR-326 in regulating invasion and migration of HCC cells.
Our results indicated that miR-326 expression decreased in HCC cells but increased in exosomes. A recent study has pointed out that miR-326 expression is declined in HCC tissues [20]. Another study has presented miR-326 expression is notably reduced in HCC cell lines and tissues and its down-regulation predicts a poor prognosis in HCC [13]. It is reported that miR-326 acts a tumor-suppression role and is greatly depressed in HCC cells [21]. All these aforementioned evidences are in line with our findings. A study has purported that in comparison with the controls, miR-326 expression is raised in Tconv-derived exosomes which is observed in relapsing–remitting multiple sclerosis patients [11].
Other results emerge from our data that highly expressed exosomal miR-326 reduced cell proliferation, colony formation, migration and invasion as well as facilitated apoptosis of HCC cells in vitro and reduced the volume and weight of HCC tumor in vivo. It has been suggested previously that HCC cell growth can be suppressed via overexpression of miR-326, and HCC cell migration and invasion ability are markedly attenuated through elevating miR-326 [21]. It is reported that the up-regulated miR-326 expression suppresses HCC cell growth and invasiveness as well as stimulates cell apoptosis in vitro [14]. Besides that, a prior study has verified that overexpression of miR-326 declined tumor growth in vivo [13]. A study has revealed that ectopic expressed miR-326 markedly attenuates cell growth, and suppresses cellular migration and invasiveness in non-small cell lung cancer cell lines [22]. Moreover, it is found that miR-326 decreases profibrotic genes like MMP-9, implying its repressive function in cancer cell proliferation [23]. Also, it is presented that miR-326 represses Bcl-2 protein expression and elevates Bax expression so as to affect the apoptosis [24]. Similar to our findings, there are some miRNAs interacting with exosomes to play a role in HCC development. It is displayed that highly expressed exosomal miR-638 can repress the proliferation of HCC cells, involving the potential impact on carcinogenesis [25]. Another study also proves that HCC cells-derived exosomal miR-451a suppresses tumor angiogenesis via disrupting endothelial functions as apoptosis, tube formation, migration and permeability [26]. A prior research generally confirms that when treated with the overexpression of miR-744 exosomes, the proliferation of HCC cells is dramatically suppressed [27].
Conclusion
To briefly conclude, our study provides evidence that M1 macrophage-derived exosomal miR-326 suppresses proliferation, migration and invasion as well as advances apoptosis of HCC cells, supplying a new insight in a novel target therapy for HCC. Due to the limited sample size and limited known researches, the exact mechanism of miR-326 is not fully elucidated, and therefore, further large-scale studies are required to illustrate the underlying mechanism.