나노기술은 빠르게 발전해 왔으며 현재 많은 첨단 의료 치료법에 사용되고 있습니다. 그러나 나노 입자(NP)에 대한 노출이 후성 유전 기전이 점점 더 많은 질병과 관련되기 때문에 다른 전신 질병을 유발할 수 있다는 우려가 증가하고 있습니다. NP 후성유전체 변형의 역할은 질병 병인에 중요합니다. 우리의 연구는 세포를 SiO2에 노출시켜 폐와 고환 세포의 손상에 대한 후성 유전적 메커니즘을 결정하는 것을 목표로 했습니다. NP. SiO2의 손상 효과를 특성화하기 위해 수컷 C57BL/6 마우스를 사용했습니다. 폐 및 고환 세포의 NP 및 각인된 Dlk1/Dio3 도메인 영역에서 결과적인 메틸화 상태. SiO2에 노출된 A549 셀 NP는 세포 사멸이 있었고 수컷 마우스는 SiO2에 노출되었습니다. NP는 폐 및 고환 조직을 변경했습니다. 각인된 도메인 Dlk1/Dio3 영역의 유전자는 두 조직 모두에서 변경되었습니다. Dlk1 , RT1 , 및 디오3Dlk1 동안 고환에서 상향 조절됩니다. 및 디오3 또한 폐 조직에서 상향 조절됩니다. 남성 성인 폐와 고환의 Bisulfite 염기서열분석 PCR은 대부분 과메틸화된 CpG와 함께 대부분 저메틸화되었습니다. 이러한 발견은 나노입자가 각인된 유전자의 DNA 메틸화에 중요한 역할을 한다는 것을 나타냅니다.
<섹션 데이터-제목="배경">
배경
이산화규소는 화학식 SiO2를 갖는 실리콘 산화물입니다. , 자연에서 석영과 다양한 유기 환경에서 가장 일반적으로 발견됩니다[1]. 가공된 나노 입자는 첨단 산업에서 나노 기술의 급속한 성장과 응용에 널리 퍼져 있습니다. 이 특정 나노 입자는 큰 비표면적, 풍부한 반응성 부위, 높은 표면 에너지, 불포화 화학 결합, 강력한 흡착 능력, 금속 및 유기 물질과 상호 작용하는 경향이 강하여 환경에서 오염 물질과 그 이동을 변경합니다[2]. SiO2의 존재 광범위한 소모품에 포함된 나노입자(NP)는 환경에 방출될 가능성을 높이고 인구와 접촉하게 됩니다.
이전의 실험 연구에서는 단일 용량의 기관 내 점적 또는 금속 및 금속 산화물 나노 입자 종의 다중 복강 주사가 세포에서 전신 및 유기체 수준까지 독성 효과를 일으키는 것으로 나타났습니다[3]. SiO2 처리 NP는 세포 사멸을 증가시키고 세포 운동성을 감소시켜 유방암 세포주의 성장을 억제합니다. 또한 SiO2에 대한 노출 나노입자는 표피성장인자수용체(EGFR)를 상당히 교란시킨다[4]. 쥐 모델이 3가지 다른 크기의 TiO2로 처리되었을 때 NPs 및 대조군과 비교하여, 큰 덩어리(> 100nm) 에어로졸을 사용한 기관지폐포 세척액(BALF) 처리는 급성 염증 반응을 유도한 반면, 작은 덩어리(<100nm) 에어로졸은 상당한 산화 스트레스 손상과 세포독성을 생성했습니다[5].
번식에 대한 나노입자 독성 연구는 성장하는 분야입니다. 한 연구에 따르면 동일한 치료 용량에서 Ni NP는 C에서 더 높은 생식 독성을 유도했습니다. 엘레건 Ni MP(미세입자)보다 이러한 생식 독성은 C에서 관찰되었습니다. 엘레건 감소된 새끼 크기, 수정란 및 정자 활성화가 포함됩니다[6]. 특정 환경적 영향이 정자 게놈의 변화 없이 부계 경로를 통해 자손에게 전달될 수 있다는 증거가 증가하고 있습니다[7, 8]. 부계 정보는 게놈뿐만 아니라 관련된 특정 후성유전학적 마커, mRNA 함량 및 비암호화 RNA에도 존재합니다.
산화 스트레스는 DNA 손상, 염증, 단백질 변성 및 지질 과산화를 유발할 수 있는 나노입자 독성의 중요한 메커니즘입니다[9]. 이러한 생물학적 효과는 크기, 표면적, 모양, 표면 화학, 기능화 및 용해도를 포함한 나노 입자의 물리화학적 특성에 영향을 받습니다[10, 11]. 나노입자에 대한 노출이 세포독성이 없는 낮은 용량에서도 조직과 세포에서 후성유전적 변화를 유발할 수 있다는 증거가 증가하고 있습니다[12, 13]. 후성유전학은 DNA의 메틸화, 유전자 각인, 히스톤 변형 및 비암호화 RNA에 의한 조절을 포함하는 DNA 서열의 변화를 포함하지 않는 유전자 기능의 유전 가능한 변화에 대한 연구입니다[14]. 이러한 후성 유전적 변화는 수많은 병리학적 상태 및 질병의 발달 및 진행과 관련이 있습니다[15]. 따라서 후성 유전적 영향은 세포 수준에서 환자 위험 평가 스크리닝의 중요한 부분입니다.
Dlk1/Dio3 각인 도메인은 부계에서 메틸화되는 3개의 알려진 차등 메틸화 영역(DMR)을 포함합니다:유전자간 DMR(IG-DMR), 모계에서 발현된 3-DMR(Gtl2-DMR) 및 Dlk1-DMR 이전 연구에서는 IG-DMR이 Gtl2의 대립형질 메틸화 상태를 지시한다고 제안합니다. 프로모터 DMR은 전체 클러스터에서 유전자 발현을 제어합니다[17]. 마우스 게놈은 12번 염색체 말단 부위의 Dlk1/Dio3 도메인에 다수의 각인된 유전자를 가지고 있습니다. 각인된 유전자 Dlk1 사이에 위치한 IG-DMR 및 Gtl2 남성 생식선에서 특이적으로 메틸화되어 각인된 유전자 영역의 부모 대립유전자 특이적 발현을 조절합니다[18]. IG-DMR 메틸화 상태는 출생 전에 확립되어 남성 생식 세포 분화 동안 남성 생식선에서 남성의 평생 동안 유지됩니다. 즉, IG-DMR 메틸화는 성숙한 고환의 정자와 정자 세포에서 유지됩니다.
우리의 목표는 환경 변화를 통해 자손에 대한 부계 영향을 설명하기 위해 전사 및 번역 침묵 전에 정자 형성 동안 남성 생식선 유전자 발현의 변화를 찾는 것이 었습니다. 환경 요인은 정자의 전사 변형을 수정할 수 있으며, 이는 자손 발달의 변화로 이어질 수 있습니다. 우리 작업에서 이 조사를 수행하기 위해 SiO2의 독성 효과를 스크리닝하기 위한 모델로 세포주와 마우스를 사용했습니다. NP. 우리가 아는 한, 이것은 나노 입자가 폐와 고환 조직 모두에 손상을 일으키는 Dlk1/Dio3 각인 영역의 후성 유전 기전을 보여주는 첫 번째 연구입니다.
방법
실험 동물
동물 취급은 Nanjing Medical University의 해당 동물 사용 프로토콜에 따라 실험 동물의 관리 및 사용에 대한 지침에 따라 수행되었습니다. 마우스는 Beijing Vital River Laboratory Animal Technology Co., Ltd.에서 구입했습니다. 모든 동물은 23°C에서 12시간 광주기로 사육되었습니다. 살균된 물과 설치류 음식은 생쥐가 마음대로 먹었습니다. 마우스 활동 및 행동을 매일 모니터링했습니다. 2주 후, 마우스에 나노 크기의 SiO2를 주입했습니다. 12.5mg/kg.
화학물질
나노 크기의 SiO2 (99.5% 미량 금속 기준, 입자 크기 10–20 nm) Sigma-Aldrich Chemical Co.(St. Louis, MO, USA)에서 입수했습니다. 나노 입자를 RPMI 1640 배지에 현탁하여 스톡 용액을 만들고 초음파 진동으로 20분 동안 분산시키고 적절한 농도로 희석한 다음 다시 20분 동안 분산시킵니다.
SiO2의 특성2 NP
크기와 제타 전위는 Malvern Zetasizer Nano ZSP를 사용하여 기록되었습니다.
RNA 추출 및 qRT-PCR
폐 및 고환 샘플을 액체 질소에서 급속 동결한 다음 약 4시간 후에 80°C에서 보관했습니다. 샘플을 추출하기 전에 샘플을 해동했습니다.
총 RNA는 1mL의 TRIzol Reagent(Invitrogen Life Technologies Co, USA)를 사용하여 샘플에서 분리되었습니다. 혼합물을 80% power에서 5분 동안 초음파 처리하고 0.2mL의 클로로포름을 첨가한 다음 12,000g에서 원심분리했습니다. 15분 동안 4°C에서 /분 그런 다음 단백질을 제거하기 위해 3단계의 페놀/클로로포름 정제를 추가했습니다. 그런 다음 UV 흡광도를 사용하여 260 및 280nm에서 각 샘플의 RNA 함량과 품질을 측정했습니다. mRNA의 프라이머 서열은 추가 파일 1:표 S1 및 S2에 나와 있습니다. qRT-PCR은 이전에 설명한 대로 제조업체의 지침을 사용하여 수행되었습니다[19]. Real-time PCR은 SYBR Green(Vazyme)을 이용하여 수행하였다. PCR 주기는 95°C에서 30초 동안 초기 변성 후 95°C에서 15초 동안 변성, 60°C에서 15초 동안 어닐링, 72°C에서 30초 확장의 40주기였습니다. 표적 유전자의 양은 β-actin으로 normalization한 후 2^-ΔCt 방법을 사용하여 분석하였다.
DNA 추출, 바이설파이트 처리 및 바이설파이트 시퀀싱 PCR
제조업체의 권장 사항에 따라 DNA 키트(QIAamp DNA Mini Kit, Qiagen. No.51304, USA)를 사용하여 고환 및 폐 조직에서 DNA를 분리했습니다. 제조업체의 권장 사항에 따라 키트(EpiTect® Bisulfite 키트, Qiagen. No. 59104, USA)를 사용하여 모든 게놈 DNA의 500ng의 바이설파이트 전환을 달성했습니다. 다른 CpG 메틸화 올리고뉴클레오티드는 Methyl Primer Express v1.0 소프트웨어를 사용하여 설계되었으며 서열은 P1-F 5'-TTGGGTTTTGAGGAGT AGTA-3', P1-R 5'-ACATCCTATTCCTAATAAAAATT-3'; P2-F 5'-TATTGGTTTGGTATATGGATGTA-3', P2-R 5'-ATAAAACACTTAACTCRTACCRTA-3'; P3-F 5'- TTTGTGTAGTTGTGTTATGGTATATTT-3', P3-R 5'- ACCCATAACAAACCACAACA-3'; P4-F 5'-TTGTGGTTTGTTATGGGTAAGTT-3', P4-R 5'-TCAAAACATTCTCCATTAACAAAA-3'.
각 DNA 샘플을 PCR에 의해 다음과 같이 증폭했습니다:2.5μl 10 × PCR 완충액 PCR 반응 믹스 500ng의 중아황산염 처리 DNA, 정방향 및 역방향 프라이머 각각 0.5μl, 0.5μl dNTP Mix, 0.5μl rTaq(500 , 마그네슘
2+
) (Takara Bio, Tokyo, Japan), ddH2 추가 O 최대 25μl의 용량. 중합효소를 94°C에서 10분 동안 활성화한 후 94°C에서 30초, 58°C에서 30초, 72°C에서 1분, 72°C에서 최종 확장의 순서로 40주기를 수행했습니다. 10분 동안 C.
세포 배양 및 치료
A549 세포는 ATCC(Manassas, VA, USA)에서 구입했으며 37°C 및 5% CO2에서 10% 소 태아 혈청(FBS)이 보충된 1640에서 배양되었습니다. 가습 인큐베이터에서. 세포를 96웰 플레이트에 플레이팅하고 다양한 농도의 SiO2와 함께 인큐베이션했습니다. NP:24시간 동안 62.5, 125, 250, 500, 1000, 2000μg/ml입니다.
세포 생존 분석
세포 생존력은 CCK8 증식 분석에 의해 평가되었습니다. 세포는 1.5 × 10
4
의 밀도로 플레이팅되었습니다. 96-well plate에 well당 넣고 밤새 배양합니다. SiO2에 노출된 후 다양한 농도의 NP, 100μl의 CCK8을 각 웰에 첨가하고 세포를 37°C에서 30분 동안 배양하여 CCK8 대사를 허용했습니다. 마침내 450nm에서 흡광도가 결정되었습니다. 세포 억제율을 계산하여 IC50로 변환했습니다. SPSS 15.0 사용.
통계 분석
모든 계산은 SPSS 15.0 소프트웨어를 사용하여 수행되었습니다. 그룹 간의 비교는 관련 없는 t를 사용하여 이루어졌습니다. 검정 및 BSP에 대한 Pearson 카이-제곱 검정. 데이터는 평균 ± SD로 표시됩니다. 모든 경우에 p 값은 <0.05는 통계적으로 유의한 것으로 간주되었습니다.
<섹션 데이터-제목="결과">
결과
SiO2의 특성2 NP
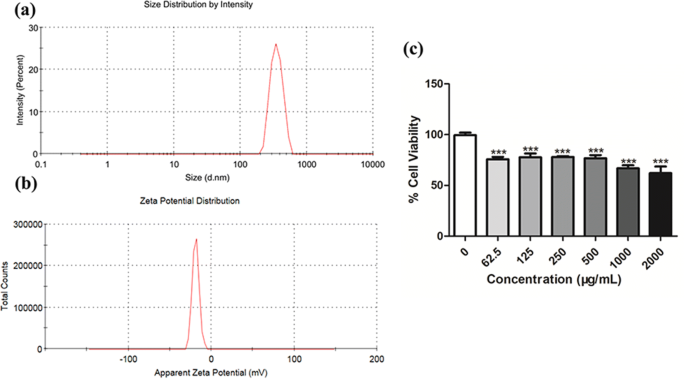
우리는 SiO2를 특성화했습니다. 실험 조건에서 NP. SiO2의 평균 유체역학적 반경 및 제타 전위 배양 배지의 NP는 각각 371.77 ± 18.46 nm 및 18.83 ± 2.12 mV였습니다(그림 1).
<그림>
SiO2의 특성화 정지된 NP. 입자를 10% FBS가 포함된 세포 배양 배지에 현탁시켰다. 아, 나 SiO2의 크기 및 제타 전위 NP는 Zetasizer Nano ZSP에 의해 평가되었습니다. ㄷ 다양한 농도의 SiO2에 노출된 후 CCK8 분석에 의해 세포 생존율을 결정했습니다. 24시간 동안의 NP 각각 3개의 기술 복제로 구성된 중복 실험의 평균 ± SD. *** p <0.001
SiO2의 효과2 A549 세포주의 NPs
SiO2의 독성을 결정하기 위해 NPs, 우리는 IC50를 결정하기 위해 A549 세포로 증식 테스트를 수행했습니다. SiO2 A549 세포의 NP. 그림 1c와 같이 SiO2 NP는 농도 의존적 방식으로 A549 세포 생존력을 감소시킵니다. 세포 생존율의 감소는 SiO2에서 현저합니다. 62.5μg/ml 이상의 NP 농도(p <0.001). IC50 24시간의 화학물질은 노출 24시간 후 세포의 50%에 영향을 미치는 농도로 정의됩니다. IC50 24시간 SiO2에 대해 결정됨 NP는 4942μg/ml였습니다.
SiO2의 효과 쥐의 폐와 고환의 NPs
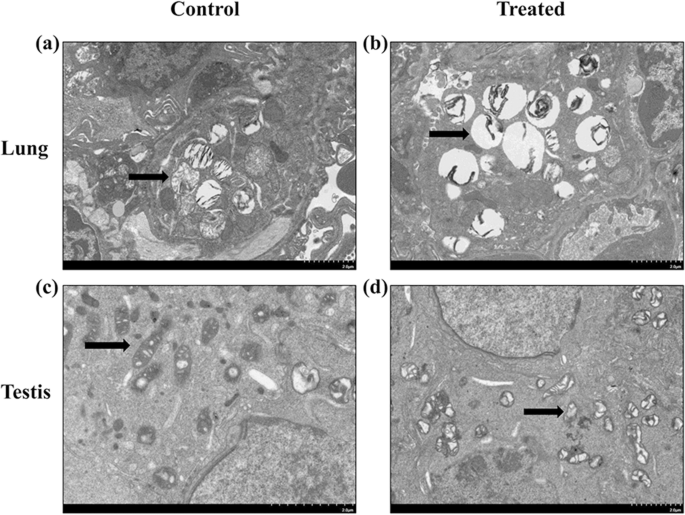
우리는 SiO2에 대한 노출 여부를 결정했습니다. 12.5mg/kg 체중의 NP는 마우스 모델에서 폐막 손상과 고환 손상을 유발할 수 있습니다. 그림과 같이 SiO2 NP 노출은 고환의 대조군과 비교하여 폐의 조직학적 섹션(그림 2a, b)과 미토콘드리아 크리스타 손상(그림 2c, d)에서 분열된 층판체를 초래했습니다. 그런 다음 SiO2의 폐 및 고환 효과를 조사했습니다. Dlk1/Dio3 각인 영역의 각인 활성화에 대한 NP.
<그림>
SiO2에 노출된 쥐의 폐 및 고환 조직의 TEM 이미지 NP. 아 형태는 대조군의 SEM에 의해 폐 조직에서 평가되었습니다. ㄴ 형태는 처리된 그룹의 SEM에 의해 폐 조직에서 평가되었습니다. ㄷ 형태는 대조군의 SEM에 의해 고환 조직에서 평가되었습니다. d 형태는 처리된 그룹에서 SEM에 의해 고환 조직에서 평가되었습니다. 스케일 바는 2.0μm
를 나타냅니다.
쥐의 폐와 고환에 각인된 유전자의 발현
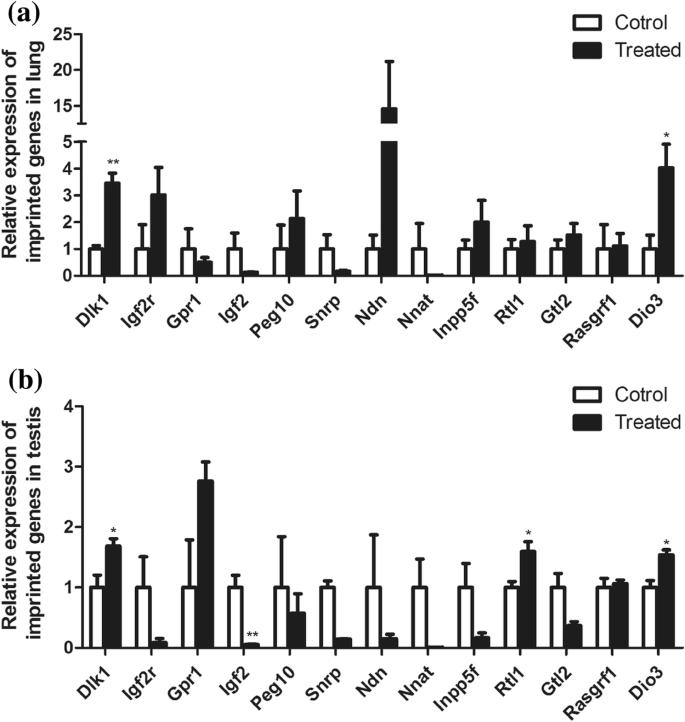
폐와 고환의 변화를 설명하기 위해 우리는 이들 조직에서 각인된 유전자를 감지했습니다. 우리는 24개의 각인된 유전자를 선택합니다. 그들은 Dio3였습니다. , Ddc , Dlk1 , Gpr1 , Gtl2 , H19 , Igf2, Igf2as , Igf2r , Inpp5f , 마겔2 , Magi2 , 메스트 , Mir296 , 미르298 , Ndn , 나트 , 페그10 , 플래그1 , Pwcr1 , Rasgrf1 , RT1 , SNRPN , 및 스너프 이 유전자 중 13개는 폐와 고환 모두에서 발현됩니다:Dio3 , Dlk1 , Gpr1 , Gtl2 , Igf2r , Igf2 , Inpp5f , Peg10, Ndn , 나트 , Rasgrf1 , RT1 , 및 Snrpn (그림 3a, b). 차등적으로 발현된 유전자는 Dlk1을 포함하는 Dlk1/Dio3 각인 영역에 일차적으로 집중되었습니다 , Gtl2 , RT1 , 및 디오3 .
<그림>
폐와 고환에서 각인된 유전자의 발현. 아 폐에서 각인된 유전자의 발현. ㄴ 고환에서 각인된 유전자의 발현. *피 <0.05. **피 <0.01. 학생의 t 테스트
Dlk1/Dio3 각인 영역의 표현
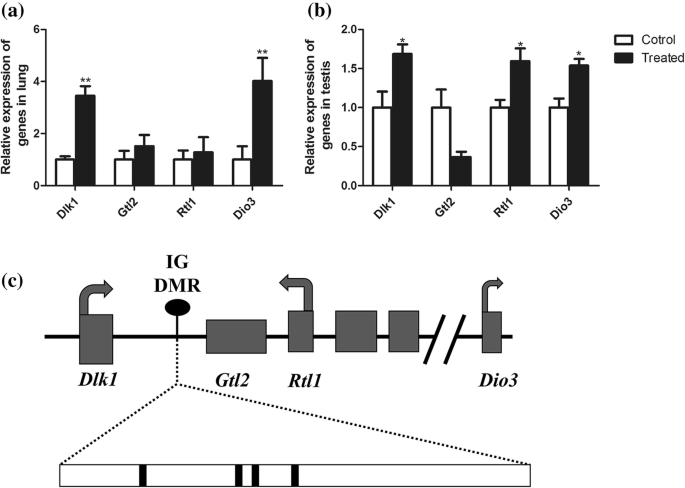
각인된 Dlk1/Dio3 영역에는 3개의 단백질 코딩 유전자(Dlk1 , Gtl2 , RT1 , 및 디오3 ) 유전된 대립유전자 [20](그림 4c). SiO2에 대한 폐 및 고환 조직 반응에서 Dlk1/Dio3 영역의 역할 설명 NP 처리, 우리는 대조군과 비교하여 DMR의 메틸화 패턴을 분석했습니다. 다른 유전자는 폐와 고환에서 메틸화의 표적이 됩니다. Dlk1의 표현 및 디오3 폐와 고환 모두에서 상향조절된 반면 Rtl1 고환에서만 상향 조절되었습니다(그림 4a, b).
<그림>
Dlk1/Dio3 각인 영역의 표현. 아 폐에서 발현되는 Dlk1/Dio3 영역 유전자. ㄴ 고환에서 발현되는 Dlk1/Dio3 영역 유전자. ㄷ Dlk1/Dio3 영역의 스키마입니다. *피 <0.05. **피 <0.01. 학생의 t 테스트
Dlk1/Dio3 DMR 영역의 메틸화
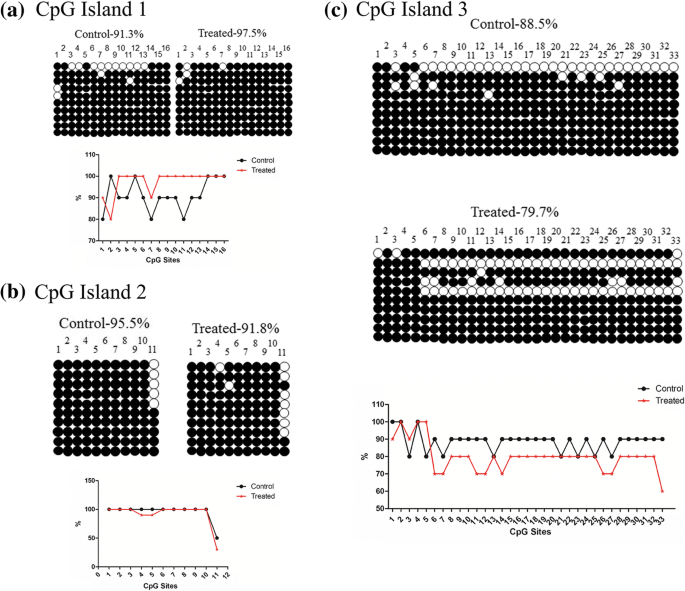
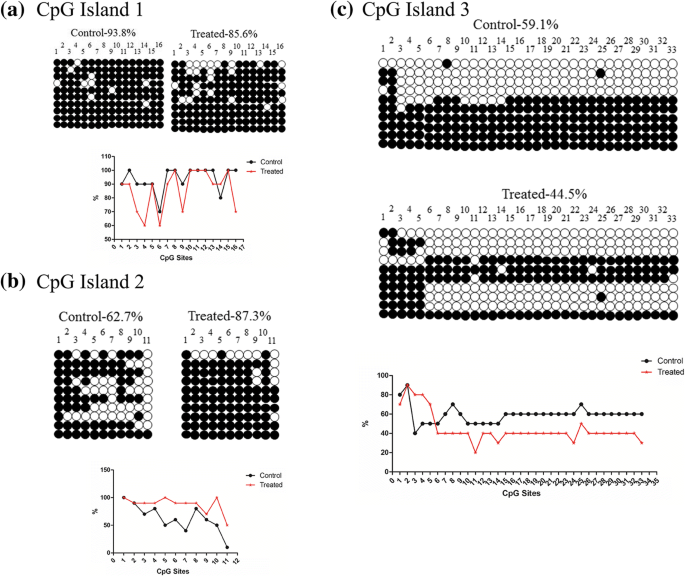
DNA 메틸화에 대한 반응으로 유전자 발현이 변화하는지 여부를 더 조사하기 위해 우리는 마우스 폐와 고환에서 이 영역의 메틸화 상태를 다루었습니다. DNA 메틸화 분석에서 우리는 CpG 섬의 세 부분의 서열을 결정했습니다. 고환에서는 저메틸화되어 있습니다. 그러나 CpG 섬 1에서는 상당히 과메틸화되어 있습니다(그림 5). 폐에서 전체 메틸화는 고환에서와 동일하지만 CpG 섬 2는 과메틸화를 나타냈다(그림 6).
<그림>
고환에서 Dlk1/Dio3 DMR 영역의 메틸화. 아 대조군 및 처리된 조직에서 CpG 섬 1의 메틸화. ㄴ 대조군 및 처리된 조직에서 CpG 섬 2의 메틸화. ㄷ 대조군 및 처리된 조직에서 CpG 섬 3의 메틸화
<그림>
폐에서 Dlk1/Dio3 DMR 영역의 메틸화. 아 대조군 및 처리된 조직에서 CpG 섬 1의 메틸화. ㄴ 대조군 및 처리된 조직에서 CpG 섬 2의 메틸화. ㄷ 대조군 및 처리된 조직에서 CpG 섬 3의 메틸화
토론
나노 물질의 사용이 증가함에 따라 인간 건강 및 환경 영향에 대한 잠재적 영향에 대한 우려가 제기되었습니다. 이전 연구에서는 SiO2가 NP는 폐 염증 및 늙은 쥐의 심근 허혈성 손상과 같은 폐 및 심혈관 손상을 유발할 수 있습니다[21]. 더욱이 나노입자는 생식계열에 영향을 미칠 수 있는데, 그 이유는 그러한 세포가 Ag 나노입자의 독성 효과에 더 민감한 것으로 보이고 더 낮은 용량에 노출된 후 부작용을 나타내기 때문입니다. Ag NP 노출은 쥐의 정자 세포에서 관찰되는 이상 수를 증가시켰고 미토콘드리아 활성을 감소시키는 것 외에도 첨체와 원형질막의 완전성을 감소시켰습니다[22]. 우리의 조사는 수컷 유기체와 노출되지 않은 자손을 표적으로 삼는 나노입자의 잠재력을 평가하기 위해 실험 플랫폼을 사용하는 일련의 연구의 일부입니다.
이전의 시험관 내 연구에서 우리는 일부 나노 입자에 대한 단기 노출이 TM-4 Sertoli 세포에서 세포 사멸 및 각인된 유전자의 비정상적인 발현을 초래한다고 보고했습니다. 이러한 발견은 각인된 유전자의 비정상적 발현이 나노입자가 생식 독성을 유발하는 기본 메커니즘일 수 있음을 보여줍니다[23]. 또한, 우리의 이전 생체 내 연구에서 내분비 교란 물질과 같은 일부 환경 요인은 노출된 개인뿐만 아니라 연속적인 자손 세대에서도 표현형 또는 질병 상태를 촉진합니다. 영구적으로 프로그래밍되는 생식계열의 후성 돌연변이는 후성 유전적 형질전환 표현형의 전달을 허용할 수 있습니다[19]. 이 연구의 목적은 SiO2에 의한 후성유전학적 상태의 변화를 조사하는 것이었습니다. 수컷 세대교체 효과의 기계적 토대를 마련하기 위한 쥐 모델의 NP 처리
후성유전학적 상태는 DNA 서열의 변화 없이 게놈 내에서 일어나는 화학적 변형을 정의하는 데 사용되는 용어이다[24]. DNA 메틸화, 각인된 유전자, 히스톤 변형 및 비암호화 RNA 발현을 포함한 후성유전학적 메커니즘은 외인성 환경에서 게놈 기능에 영향을 미칠 수 있습니다[25]. 우리가 아는 한, 우리 연구는 SiO2를 조사한 첫 번째 연구입니다. 후성적 수준에서 폐 및 고환 독성을 유도하는 NP.
우리는 먼저 SiO2의 급성 독성을 조사했습니다. 인간 폐 상피 세포주인 A549 세포의 NP. 그러나 실험 쥐에 대한 우리의 발견은 SiO2와 접촉한 후 층류 폐 유형 II 상피 세포의 손상과 고환 미토콘드리아 능선 손상을 보여주었습니다. 환경 농도의 NP [26]. 폐 및 고환 병리의 메커니즘을 더 잘 이해하기 위해 각인된 유전자를 표현했습니다. 게놈 각인은 기원의 부모에 따라 배우자의 접합체에서 한 부모 대립 유전자의 침묵을 나타냅니다. 이 침묵은 DNA 메틸화 및/또는 히스톤 변형과 같은 후생적 과정을 통해 발생합니다[27]. 이전 연구에서는 Dlk1/Dio3 도메인의 각인된 유전자 발현이 태아 성장[28], 인간 사춘기의 시기[29], 대사 질환에 대한 감수성[30]에 중요하다는 것을 보여주었습니다. 연구에 따르면 IG-DMR은 Gtl2 프로모터 DMR의 대립형질 메틸화 상태를 지시하며, 이는 전체 Dlk1/Dio3 영역에 걸쳐 유전자 발현을 제어합니다[17]. 이 각인된 제어 영역의 주요 기능은 생식 세포에 의한 DNA 메틸화를 배우자 신호로 상속하고 나중에 체세포 내에서 후속 대립유전자 특이적 DNA 메틸화 패턴을 유지하는 것입니다[31]. 우리의 연구는 SiO2 NP는 폐 및 고환 조직 모두에서 Dlk1/Dio3 영역의 발현에 변화를 유도합니다. Dlk1/Dio3 영역에서 부계 발현 유전자(Dlk1 , RT1 , 및 디오3 )는 NP 처리 후 대조군에 비해 특히 비정상적이다. Bisulfite 시퀀싱 결과는 폐와 고환에서 다양한 수준의 저메틸화를 나타냅니다. IG-DMR의 메틸화 상태는 일반적으로 처리된 조직에서 더 낮으며, 이러한 저메틸화는 각인된 유전자의 차등 발현 메커니즘을 나타낼 수 있습니다.
결론
결론적으로, 우리의 결과는 SiO2 NP 노출은 세포 손상을 유발하는 중요한 DNA 메틸화 변화를 유도할 수 있으며 이러한 변화는 Dlk1/Dio3 각인 유전자 클러스터의 발현에 매우 중요합니다. 중요한 것은 DNA 메틸화의 변화가 폐와 고환 조직 모두에 영향을 미친다는 것입니다. 이러한 결과는 노출된 모델의 자손에 의해 유전되는 나노입자의 후성유전학적 효과를 조사하고 그러한 후성유전적 변화를 매개하는 분자 메커니즘의 설명을 조사하는 우리의 미래 연구에서 중요한 역할을 합니다.