금 나노입자 억제 시토크롬 P450 3A4 활성 및 인간 간세포 암종 세포주 C3A에서 세포 독성의 기초가 되는 분자 메커니즘의 평가
초록
양이온성 분지형 폴리에틸렌이민(BPEI), 음이온성 리포산(LA) 또는 중성 폴리에틸렌 글리콜(PEG)로 기능화된 40 및 80nm 금 나노입자(AuNP)와 인간 간세포 암종(HCC) 세포주 C3A의 상호 작용이 조사되었습니다. 인간 혈장 단백질 코로나(PC)의 부재 및 존재. 80nm LA-AuNP를 제외한 모든 베어(PC 없음) AuNP는 C3A에 세포독성이 있었지만 PC는 세포독성을 약화시켰습니다. AuNP의 시간 의존적 세포 흡수는 40nm BPEI-AuNP 외에 증가했지만 PC는 80nm PEG-AuNP 외에는 흡수를 억제했습니다. BPEI-AuNP에 의한 산화/질산화 스트레스의 2상 반응은 C3A 세포에서 발생한 반면, PEG-AuNP는 강력한 항산화제였습니다. 모든 베어 AuNP는 크기 및 표면 전하에 관계없이 시토크롬 P450(CYP) 3A4 활성을 억제했지만 PC는 PEG-AuNP 외에 활성을 회복했습니다. 40nm PEG-AuNP 조절 유전자 발현은 주로 미토콘드리아 지방산 β-산화 및 간 유출/흡수 수송체와 관련이 있습니다. 이러한 연구는 AuNP가 주요 생물학적 과정과 HCC의 기본 분자 메커니즘과 상호작용하는 것을 더 잘 이해하는 데 기여하며, 이는 HCC 치료에서 보다 효과적인 치료 표적의 개발에 추가로 연루될 수 있습니다.
<섹션 데이터-제목="배경">
배경
간세포암종(HCC)은 전 세계적으로 가장 흔한 암 중 하나이며 미국에서 암 사망률의 가장 빠르게 증가하는 원인입니다[1, 2]. 간세포암종은 이미 진행된 단계에서 진단되었기 때문에 간세포암종 치료는 종양 발달 초기에 간이식 또는 외과적 절제술과 종양이 진행된 상태에 대한 화학 요법 및 방사선 요법이 있습니다. HCC는 종종 전신적 부작용을 초래할 수 있는 비선택적 세포독성 분자인 기존의 항종양제에 대해 높은 내성을 나타냅니다. 최근 유전자 치료의 발전, 즉 RNA 간섭(RNAi) 기반 유전자 치료가 현재 간세포암종 치료에 활용되고 있다[3, 4]. RNAi의 효능은 벡터가 표적 세포의 내부로 전달되어야 한다[5]. 성공적인 유전자 전달을 위한 벡터는 바이러스 및 비바이러스 벡터입니다. 바이러스는 유전자 전달의 효율성이 더 높지만 바이러스 벡터의 안전성 문제로 인해 비바이러스 벡터가 선호됩니다. 표적 유전자 전달 또는 약물 전달 시스템을 위한 비바이러스 벡터로서의 나노입자(NP)는 HCC 치료에서 치료 효율을 개선하고 전신 및/또는 세포 수준에 대한 독성을 낮추는 데 큰 주목을 받고 있습니다[4, 6]. 따라서 표적 세포 및 조직에서 NP의 세포 교란 및 독성의 기저에 있는 분자 메커니즘 및 생물학적 경로를 확인하는 것이 매우 중요합니다. 최근의 시험관 내 연구는 세포 및 생화학적 반응과 결합된 유전자 발현 프로파일링이 세포 교란 및 잠재적인 NP 독성에 대한 직접적인 평가를 제공한다는 것을 보여주었습니다[7,8,9,10].
금 나노입자(AuNP)는 독특한 물리화학적 특성 및 표면 화학으로 인해 단독으로 또는 다른 약물과 함께 유전자 침묵 부분의 표적 특이적 전달을 위한 전달 수단으로 사용되었습니다[11, 12]. AuNP와 혈장 단백질의 상호작용은 단백질 코로나를 형성하고, 이는 차례로 NP 표면 화학을 변경하고 세포 흡수 및 잠재적 독성과 같은 후속 생물학적 반응에 영향을 미칩니다[13, 14]. 다양한 인간 암세포주와 1차 세포에서 AuNP의 세포 흡수는 크기와 표면 전하에 관계없이 단백질 코로나 형성에 의해 결정적으로 영향을 받았습니다[7,8,9, 14,15,16,17].
크기 및 표면 전하 의존적 산화 스트레스는 NP 세포 독성과 관련된 AuNP에 반응하여 인간 유방암 세포주, MDA-MB-231, 간세포 암종 HepG2 및 인간 백혈병 HL-60 세포에서도 관찰되었습니다 [18 , 19]. AuNP에 의한 세포독성은 다양한 인간 암세포주와 1차 인간 세포에서 세포 유형 특이적 방식으로 발생하였다[7,8,9,20,21].
Cytochrome P450(CYP) 효소는 수많은 세포독성 약물의 생물학적 활성화 또는 비활성화와 항암제의 발암성에 대한 숙주 감수성에 중요한 역할을 합니다[22]. AuNP는 생체 내 및 시험관 내 세포 및 분자 수준에서 CYP 효소의 촉매 활성에 영향을 미쳤습니다[7, 23, 24, 25]. AuNP는 인간 폐 섬유아세포 세포주 MRC-5의 산화 스트레스 마커에 주로 관여하는 차등 유전자 발현과 인간 제대 정맥 세포(HUVEC) 및 인간 간세포의 미토콘드리아 기능 장애를 상당히 나타내었으며, 이는 과산화지질 생성 및 높은 세포 독성 [8, 9, 26]. 이 지식은 AuNP가 다양한 세포 유형에서 세포 사멸 또는 괴사 세포 사멸을 유발하고 스트레스 반응 경로 및 독성에서 차등적인 유전자 발현과 결합된 세포 및 생화학적 기능을 변경한다는 것을 상호 제안하지만 AuNP가 세포 또는 생물학적 내에서 독성 효과를 발휘하는 특정 경로를 통해 시스템은 알 수 없는 상태로 남아 있습니다.
여기에서 이 연구는 인간 HCC 세포 C3A와 AuNP 상호작용에 대한 단백질 코로나, 크기 및 표면 전하의 영향을 조사했습니다. 주로 C3A 세포에서 양이온성 BPEI, 음이온성 리포산(LA) 또는 중성 폴리에틸렌 글리콜(PEG)로 기능화된 40 및 80nm AuNP의 시간 의존적 세포 흡수가 인간 혈장 단백질 코로나(PC) 유무에 따라 결정되었습니다. 둘째, AuNP 유도 세포독성 및 반응성 산소종(ROS)/반응성 질소종(RNS) 생성을 CYP3A4 활성에 대한 억제 효과와 함께 모니터링했습니다. 마지막으로 AuNP 독성 관련 분자 작용 메커니즘은 Human Molecular Toxicology Pathway Finder 및 Human Drug Transporters RT
2
를 사용하여 특성화되었습니다. Profiler™ PCR 어레이.
방법
금 나노입자 합성
40 및 80nm 양이온 BPEI, 음이온 LA 및 중성 PEG Biopure™ AuNP는 nanoComposix(San Diego, CA)에서 맞춤 합성되었습니다. 입자 크기, 다분산 지수(PDI), 제타(z) 전위 및 스펙트럼 특성은 동적 광산란(DLS), 투과 전자 현미경(TEM) 및 UV-Vis 분광법으로 특성화되었습니다. AuNP는 탄산칼륨 수용액에서 사염화금수소(III) 수화물을 환원시킨 후 숙성 과정과 접선 유동 여과(TFF)를 통해 합성되었습니다. AuNP 표면은 디하이드로리포산(0.2:1, w /와 ) 또는 티올-메톡시-말단 PEG(Laysan Bio Inc., Arab, AL)(0.5:1, w /와 ), 각각 TFF 세척 및 멸균 여과가 뒤따릅니다. AuNP의 BPEI 기능화된 표면은 LA의 카르복실산을 BPEI의 유리 아민에 연결한 다음 TFF 세척 및 결합되지 않은 BPEI 제거를 위한 원심분리를 통해 EDC 화학을 통해 합성되었습니다.
단백질 코로나 준비
합동 인간 혈장(n =5) Biological Specialty Corp.(Colmar, PA)에서 입수했습니다. AuNP는 인간 혈장에서 배양되었습니다(55%, v /v ) 보고된 대로 1시간 동안 37°C에서 250rpm의 일정한 속도로 작동합니다[7, 8]. 결합되지 않은 약하게 결합된 단백질은 20,000xg에서 인산염 완충 식염수(PBS)로 반복 세척하여 제거했습니다. 20°C에서 20분 동안 최종 인간 혈장 단백질 코로나(PC) 코팅된 AuNP를 PBS에 분산시킨 다음 추가 물리화학적 특성화 또는 투여를 위해 세포 배양 배지에 희석하였다. 자세한 프로토콜은 추가 파일 1에 나와 있습니다.
AuNP의 물리화학적 특성
유체역학적 직경(DH ), PDI, 탈이온수(DI)에서 BPEI, LA 및 PEG로 기능화된 40 및 80nm 베어(PC 없음) AuNP의 z-전위는 Zetasizer Nano ZS(Malvern)를 사용하여 0시간에 25°C에서 분석되었습니다. Instruments, Worcestershire, UK); PBS에서 PC 코팅된 AuNP의 경우 25°C, 0시간; 0시간 및 24시간에 37°C에서 완전한 세포 배양 배지의 모든 베어 및 PC AuNP에 대해. 완전한 세포 배양 배지에는 10% FBS(ATCC
®
)가 보충된 Eagle의 최소 필수 배지(EMEM)가 포함되어 있습니다. , 매나사스, 버지니아). 샘플은 각각 10초씩 11개의 하위 실행으로 5회 측정되었습니다. 또한, 광학 흡수 스펙트럼은 Synergy H1 하이브리드 다중 모드 마이크로플레이트 판독기(BioTek Instruments Inc., Winooski, VT)를 사용하여 실온 0시간에서 측정되었습니다.
투과 전자 현미경
AuNP 형태는 TEM을 사용하여 특성화되었습니다. 모든 베어 및 PC AuNP 용액(5μL)을 200메시 구리 그리드에 놓고 실온에서 공기 건조했습니다. 샘플은 120kV의 가속 전압에서 Oxford 검출기(FEI Company, Hillsboro, OR)가 있는 Tecnai G2 Spirit BioTWIN에서 관찰되었습니다. GATAN 현미경 검사 세트(GATAN Inc., Pleasanton, CA)는 AuNP 직경을 측정했습니다.
세포 배양 및 생존력 측정
인간 간세포 암종 C3A 세포(ATCC
®
CRL-10741™) ATCC
®
에서 구입했습니다. (Manassas, VA), 완전한 EMEM에서 배양(ATCC
®
, Manassas, VA) 10% FBS로 보충하고 4일마다 배지를 교체하면서 T75 플라스크에서 약 80% 합류로 확장했습니다. 0.25% 이후(w /v ) 트립신-0.53mM 에틸렌디아민테트라아세트산(EDTA) 소화, 세포를 8 × 10
4
에서 96웰 플레이트에 플레이팅 웰당 세포 및 95% 공기 및 5% CO2의 습한 분위기에서 37°C에서 배양합니다. 48시간 인큐베이션 후, PC의 부재 및 존재하에 세포에 AuNP를 투여하였다. 계대 9와 12 사이의 C3A 세포를 투여에 사용했습니다.
C3A 생존 가능성은 alamarBlue
®
를 사용하여 결정되었습니다. 생존력 분석(Thermo Sci., Waltham, MA) [7, 27]에 설명된 대로. 96웰 플레이트의 세포를 40 및 80nm BPEI-, LA- 및 PEG-AuNP로 처리하고 PC는 0~250μg/cm
2
범위에서 처리했습니다. . 24시간 후 alamarBlue
®
의 10% 완전한 EMEM의 시약(v /v )을 세포 배양에 첨가하고 37°C에서 3시간 동안 인큐베이션했습니다. 완전한 EMEM은 분산제 역할을 했습니다. AuNP와 alamarBlue
®
의 활성 성분의 상호 작용 시약, 레자주린 또는 환원 생성물, 레소루핀을 대조군으로 측정하였다. AuNP 및 레자주린(세포 없음) 또는 유지 배지(세포 없음)는 배경 대조군으로 사용되었습니다. 세포 생존율에 비례하는 형광은 대조군에 대해 정규화되었고 대조군 세포군에 대한 백분율로 표현되었습니다.
유도 결합 플라즈마 질량 분석기를 사용한 세포 흡수 측정
세포는 8 × 10
4
로 시딩되었습니다. 96웰 플레이트의 웰당 세포 및 1.56μg/cm
2
의 무독성 농도로 투여 0.5, 1, 3, 6, 12, 24시간 동안 모든 베어 및 PC AuNP의 에칭 단계는 이전에 보고된 바와 같이 세포막에 결합된 AuNP 및 웰에 대한 비특이적 결합을 제거하기 위해 통합되었습니다[28]. 세포 수확물을 왕수에서 건조 및 소화하고 NexION
™
을 사용하여 세포 내 Au 농도를 정량화했습니다. 350X 유도 결합 플라즈마 질량 분석기(ICP-MS)(PerkinElmer, Waltham, MA). AuNP의 세포 흡수는 이전에 보고된 대로 계산되었고 세포당 AuNP의 수로 표시되었습니다[29]. 자세한 프로토콜은 추가 파일 1에 나와 있습니다.
산화/질산화 스트레스 측정
세포는 8 × 10
4
로 시딩되었습니다. 96웰 플레이트의 웰당 세포 및 최대 125μg/cm
2
의 40nm 베어 BPEI 및 PEG-AuNP 투여 1, 3, 24시간 동안 산소/질산화 스트레스의 직접 측정은 이전에 설명한 대로 총 활성 산소종(ROS)/과산화물(SO) 분석 키트(Enzo Life Sciences, Farmingdale, NY)를 사용하여 분석되었습니다. ROS/반응성 질소 종(RNS)(Ex488/Em520 nm) 또는 SO(Ex550/Em610 nm)의 증가에 비례하는 형광을 마이크로플레이트 판독기로 측정했습니다. 자세한 프로토콜은 추가 파일 1에 나와 있습니다.
사이토크롬 P450 3A4 활동
CYP3A4 활성에 대한 40 및 80nm 베어 및 PC AuNP의 부작용은 완전히 설명된 대로 P450-Glo™ 분석(Promega Corp., Madison, WI)을 사용하여 특성화되었습니다. 96웰 플레이트의 C3A 세포에 중앙 치사 농도(LC50 ) 값:127.3μg/cm
2
40nm BPEI-AuNP 중 205.5μg/cm
2
80nm BPEI-AuNP의 192.5μg/cm
2
40nm LA-AuNP 및 129.5μg/cm
2
40nm PEG-AuNP의 LC50 이후 80 nm LA- 및 PEG-AuNP 값은 결정되지 않았으며 세포는 LC50로 처리되었습니다. 40nm LA- 및 PEG-AuNP 값(192.5μg/cm
2
및 129.5μg/cm
2
, 각각). 24시간 배양이 끝난 후 세포를 CYP3A4 기질(루시페린-IPA)과 함께 37°C에서 3시간 동안 배양했습니다. 효소 활성에 비례하는 발광 신호를 마이크로플레이트 판독기로 측정한 다음 대조군으로 정규화했습니다. AuNP와 모 기질 또는 대사 산물 및 무세포 기질과의 상호 작용을 평가하기 위해 대조군을 지정했습니다. CYP 활성은 대조군 세포군에 대한 백분율로 표시되었습니다.
유전자 발현 프로파일링
독성 40nm PEG-AuNP가 높은 세포 흡수를 나타내는 C3A 세포에서 CYP3A4 활성 및 항산화 활성 억제에 사용되었기 때문에 독성 및 차등적인 세포 반응의 기초가 되는 분자적 작용 메커니즘을 특성화하기 위해 선택되었습니다. 세포를 2.5 × 10
6
으로 시딩했습니다. 6웰 플레이트의 웰당 세포 및 LC50 투여 37°C에서 24시간 동안의 40nm PEG-AuNP 값 인큐베이션이 끝나면 세포를 RNA 분리한 다음 평균 RNA 무결성 번호(RIN) 값이 7.8인 총 RNA를 사용하여 cDNA 합성을 이전에 설명한 대로 수행했습니다[7,8,9]. 생성된 cDNA를 RT
2
와 혼합했습니다. SYBR 그린 마스터 믹스(Qiagen Inc., Valencia, CA) 후 Human Molecular Toxicology Pathway Finder 또는 Human Drug Transporters RT
2
에 적용 Quantstudio™ 7 Flex의 Profiler™ PCR 어레이(Applied BioSystem, Foster City, CA). 배수 변화가 <− 2 및> 2이고 p인 차별적으로 발현된 유전자 <0.05는 관심 유전자의 하향 및 상향 조절을 나타냅니다. RT
2
를 확인하려면 PCR 어레이 데이터, 선택된 9개 유전자의 발현을 cDNA 합성 및 후속 실시간 PCR로 평가하였다. 프라이머 서열은 추가 파일 1:표 S1에 요약되어 있습니다. 모든 PCR 반응은 삼중으로 수행되었습니다. Real-Time PCR 조건 및 정량에 대한 자세한 프로토콜은 추가 파일 1에 나와 있습니다.
통계 분석
치사 농도 중앙값(LC50 ) C3A 세포의 AuNP 값은 GraphPad Prism 6(La Jolla, CA)을 사용하여 관찰된 데이터(AuNP 농도 수준의 입력 및 해당 세포 생존율)에 가변 기울기가 있는 Hill 방정식을 피팅하여 추정되었습니다[7]. AuNP가 C3A 세포에서 ROS/RNS 생산 및 세포 흡수에 미치는 영향을 평가하기 위해 SAS 9.4(SAS Institute, Cary, NC)를 사용하여 일원 분산 분석(ANOVA)을 수행했습니다. 유의미한 경우 다중 비교는 p에서 Tukey의 정직한 유의차(HSD) 테스트로 수행되었습니다. <0.05.
결과 및 토론
베어 및 인간 혈장 PC AuNP의 물리화학적 특성화
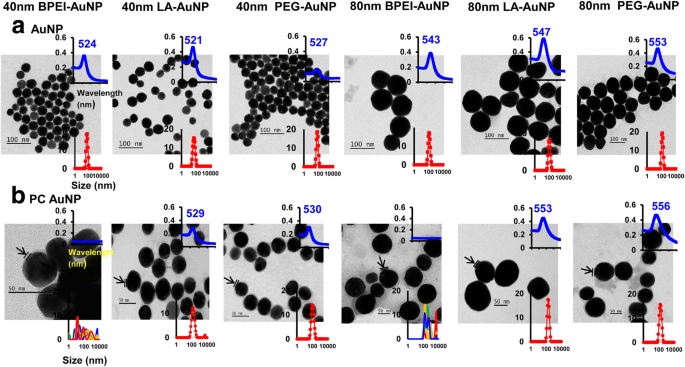
AuNP 주변의 NP 크기, 표면 전하 및 인간 플라즈마 PC 형성이 유체역학적 직경에 미치는 영향(DH ), 다분산 지수(PDI), z-전위, 스펙트럼 특성 및 형태는 DLS, TEM 및 UV-Vis 분광법을 사용하여 특성화되었습니다(그림 1). TEM 이미지에서 DI 물의 모든 베어(PC 없음) AuNP는 좁은 크기 분포와 521-553nm의 고유한 UV-Vis 스펙트럼 범위로 단분산되었습니다(그림 1a). AuNP 주변의 PC 형성은 흡수 스펙트럼의 크기 분포 및 적색 편이의 변화로 관찰되었습니다(그림 1b). 디H 완전한 EMEM에서 40 및 80nm 베어 및 PC AuNP의 PDI 값은 PDI 값이 감소한 40 및 80nm PC BPEI-AuNP를 제외하고 37°C에서 최대 24시간까지 호환 가능했습니다(각각 0.29 및 0.32 ) 37°C에서 24시간과 37°C에서 0시간(각각 0.62 및 1.0)(표 1). 모든 베어 및 PC AuNP의 Z-전위 값은 37°C에서 0시간에 비해 37°C에서 24시간에서 상대적으로 감소했습니다. PBS 및 완전한 EMEM에서 40 및 80nm PC BPEI-AuNP의 응집이 관찰되었으며, 이는 크기 분포의 다중 피크 및 DH의 변화와 상관관계가 있었습니다. 및 노출되지 않은 BPEI-AuNP에 대한 흡광도 스펙트럼의 적색 편이(그림 1 및 추가 파일 1:그림 S1, 표 1). 이러한 결과는 40 및 80 nm PC와 인간 혈청 알부민 코로나 코팅된 BPEI-AuNP가 PBS 및 다양한 세포 배양 배지에서 응집된다는 최근 연구에 의해 뒷받침됩니다[7,8,9].
<그림><그림>
a의 투과 전자 현미경 사진 탈이온수의 AuNP 및 b PBS의 PC AuNP(25°C, UV-Vis 스펙트럼 파장(상단 삽입), 동적 광산란 분포(하단 삽입)) 화살표는 PC 형성을 나타냅니다. PC 인간 혈장 단백질 코로나, BPEI 분지형 폴리에틸렌이민, LA 리포산, PEG 폴리에틸렌 글리콜
<그림>
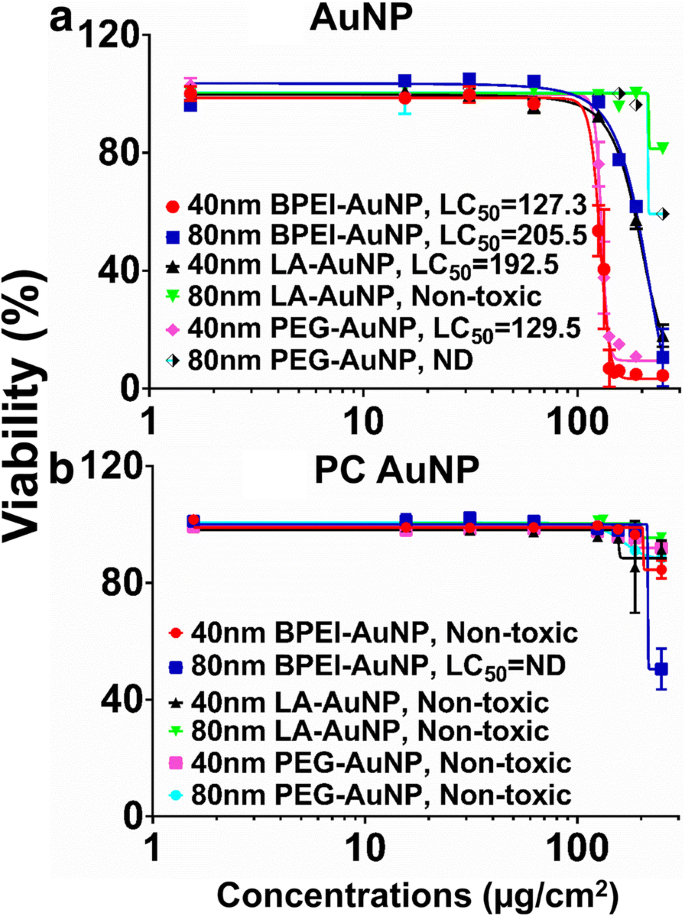
AuNP 세포독성
AuNP 세포독성은 치사 농도 중앙값(LC50 ) C3A 세포에서. NP 의존성 LC50 주변의 NP 표면 전하, 입자 크기 및 PC 형성 AuNP를 사용한 분석은 그림 2에 나와 있습니다. 모든 40nm BPEI-, LA, PEG-AuNP와 80nm BPEI-AuNP는 해당하는 LC50를 갖는 C3A 세포에 세포독성이 있었습니다. 127.3~205.5μg/cm
2
범위 (그림 2a). 80nm 베어 PEG-AuNP는 250μg/cm
2
의 최고 농도에서 59%의 세포 생존율을 나타냈습니다. , 80nm LA-AuNP는 세포독성이 없었습니다. PC는 250μg/cm2에서 51% 세포 생존율을 보인 80nm BPEI-AuNP를 제외하고 크기 및 표면 전하 변형의 함수로 AuNP 독성을 감소시켰습니다.
2
24시간에(그림 2b) 최근 연구에 따르면 40nm 베어 BPEI-AuNP가 1차 인간 간세포, HUVEC 및 인간 신장 근위 세뇨관 세포(HPTC)에 독성을 나타냅니다(LC50 22.4–80.3μg/cm
2
범위 ) [7,8,9]. PC로 코팅된 BPEI-AuNP는 인간 간세포에 세포독성을 나타내었지만 HSA로 코팅된 AuNP는 세포독성이 없었다[7]. 이러한 결과는 C3A 세포가 암 세포주의 높은 증식 속도와 대사 활성으로 인해 1차 인간 세포보다 AuNP 독성에 더 내성이 있음을 시사한다[31].
<그림><그림>
C3A 실행 가능성 및 LC50 40 및 80nm a 값 AuNP 및 b PC AuNP. 데이터는 평균 ± S.D.를 나타냅니다. (n =3). PC 인간 혈장 단백질 코로나, ND 미결정, BPEI 분지형 폴리에틸렌이민, LA 리포산, PEG 폴리에틸렌 글리콜, LC50 치사 농도 중앙값
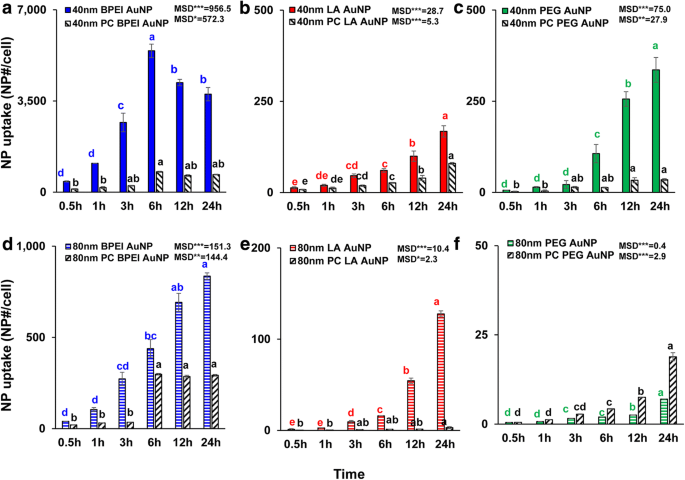
AuNP의 세포내 흡수
모든 베어 및 PC AuNP의 NP 크기, 표면 전하 및 PC 의존적 세포 흡수는 1.56μg/cm
2
에서 측정되었습니다. 최대 24시간 ANOVA는 크기, PC 및 시간에 따라 상당한 변화를 보였습니다(p <0.0001) 및 상호 작용(PC × 크기, PC × 시간, 크기 × 시간 및 PC × 크기 × 시간)(p <0.001) LA- 및 PEG-AuNP 흡수에 대한 미미한 상호 작용(PC × 크기)을 제외한 모든 AuNP 흡수(p =0.2). 그림 3a-f에서 볼 수 있듯이 40nm 및 PC AuNP 외에 40nm 및 80nm 베어 및 PC AuNP의 세포 흡수가 선형 증가하는 것이 관찰되었으며, 이는 6시간에 가장 높은 세포 흡수에 도달하고 이후 감소했습니다( 그림 3a). 그러나 24시간에 40nm 양이온성 BPEI-AuNP가 가장 높은 흡수율을 보였고 중성 40nm PEG-AuNP, 음이온성 40nm LA-AuNP가 그 뒤를 이었습니다. 이는 AuNP의 C3A 세포 세포 독성 순서와 관련이 있습니다(그림 2a). ). 이 결과는 양이온성 폴리(N-(2-아미노에틸)아크릴아미드)- 및 BPEI-AuNP가 음이온성 폴리(아크릴산)- 및 LA-AuNP 및 중성 폴리( 인간 결장직장 선암종 Caco-2 세포, HPTC 및 인간 간세포에서 N-(2,3-dihydroxypropyl)acrylamide- 및 PEG-AuNP [9, 32]. 또한 NP-PC 복합체는 40 및 80nm BPEI- C3A 세포에서 LA-AuNP 및 40nm PEG-AuNP가 있지만 80nm PEG-AuNP 흡수를 가속화했습니다(그림 3f). 이러한 결과는 PC가 HUVEC, HEK 및 HPTC에서 AuNP 흡수를 억제한다는 최근 연구에 의해 뒷받침됩니다. 크기와 표면 전하에 관계없이 [8, 9, 33] 대조적으로, PC 및 HSA 코로나는 인간 간세포에서 40nm PEG-AuNP 흡수를 향상시켰지만 후자는 HEK에서 80nm PEG-AuNP 흡수를 유도했습니다[7, 33] .
<그림><그림>
40nm a의 시간 의존적 세포 흡수 BPEI-AuNP, b LA-AuNP 및 c PEG-AuNP 및 80nm d BPEI-AuNP, e LA-AuNP 및 f 최대 24시간 동안 C3A 세포에서 PC의 부재 및 존재에서 PEG-AuNP 데이터는 평균 ± S.D.를 나타냅니다. (n =3). Tukey의 HSD 테스트에 따라 문자가 크게 달랐습니다. BPEI 분지형 폴리에틸렌이민, LA 리포산, PEG 폴리에틸렌 글리콜, PC 인간 혈장 단백질 코로나, MSD 최소 유의차. *p <0.05; ** p <0.005; ***p <0.0001
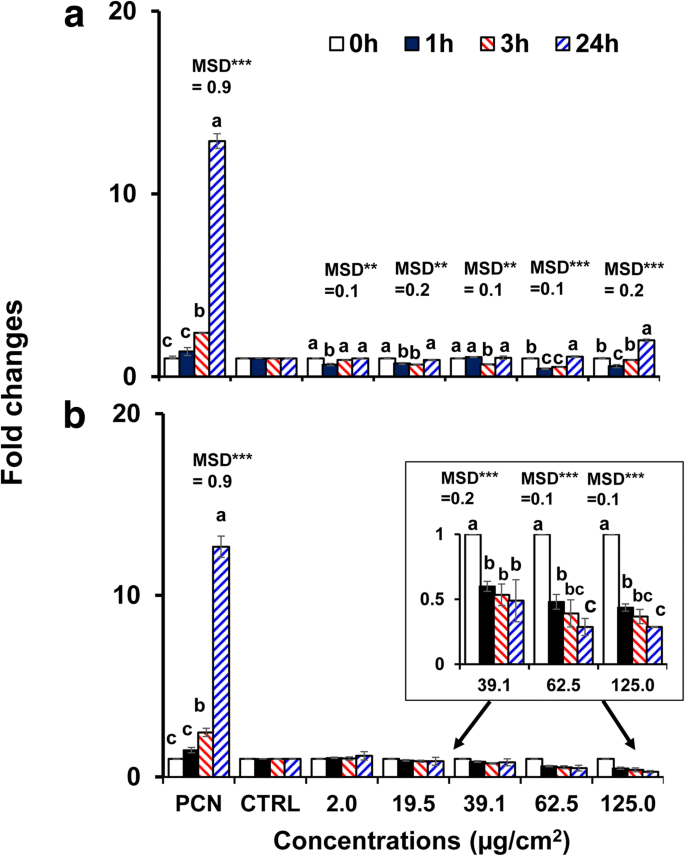
산화 및 질산화 스트레스 측정
40nm 베어 BPEI- 및 PEG-AuNP는 다른 AuNP에 비해 C3A 세포에서 더 높은 세포독성 및 세포 흡수를 나타내므로 AuNP 유도 산화/질산화 스트레스를 조사하기 위해 선택되었습니다. AuNP는 C3A 세포에서 시간 및 농도 의존적 방식으로 ROS/RNS 생성을 조절했습니다(p <0.0001) 및 상호 작용(시간 × 농도, p <0.0001). 그림 4a에서 볼 수 있듯이 ROS/RNS 생성은 40nm BPEI-AuNP(62.5μg/cm
2
)의 농도가 높을수록 감소했습니다. 및 125μg/cm
2
) 37°C에서 1시간, 24시간까지 증가했습니다. 대조적으로, 40nm PEG-AuNP는 39.1μg/cm
2
에서 ROS/RNS 생성을 실질적으로 억제했습니다. 최대 24시간까지 배수 변화 <0.5로 계속 진행합니다(그림 4b). 세포 사멸의 활성화는 종종 NP 독성에 기여하며 대부분의 경우 산화 스트레스로 이어지는 ROS/RNS 생성의 증가가 NP 독성의 원인이 됩니다[34]. 표면 전하 의존적 ROS/RNS 생성은 40nm 양이온성 BPEI- 및 중성 PEG-AuNP에서 관찰되었습니다. 40nm BPEI-AuNP는 고농도에서 ROS/RNS 생성의 2상 패턴(1시간에 항산화제, 3시간 이후에 산화촉진제)을 보여주었으며, 이는 C3A 세포에서의 세포독성과 관련이 있었습니다(그림 2a). 이 결과는 40nm 및 80nm BPEI-AuNP 및 20nm 시트레이트-AuNP 유도 ROS 생성이 각각 시간 및 농도 의존적 방식으로 인간 간세포 및 HepG2 세포의 세포독성과 관련이 있다는 이전 연구와 일치합니다. [7, 35]. AuNP는 크기에 관계없이 인간 전골수구성 백혈병 세포인 HL-60에서 총 글루타티온 감소와 함께 산화 스트레스 유발 세포독성을 보였다[19]. 대조적으로, 40nm PEG-AuNP는 산화/질산화 스트레스가 C3A 세포에서 40nm PEG-AuNP 유도 세포독성의 직접적인 메커니즘이 아닐 수 있음을 시사하는 항산화제 역할을 했습니다(그림 2b).
<그림><그림>
a에 노출된 C3A 세포에서 시간 및 농도 의존적 ROS/RNS 생성 40nm BPEI-AuNP 및 b 40nm PEG-AuNP 최대 24시간 데이터는 평균 ± S.D.를 나타냅니다. (n =3). Tukey의 HSD 테스트에 따라 문자가 크게 달랐습니다. BPEI 분지형 폴리에틸렌이민, LA 리포산, PEG 폴리에틸렌 글리콜, CTRL 제어, MSD 최소 유의차, PCN 피오시아닌(ROS 유도제). ** p <0.005; ***p <0.0001
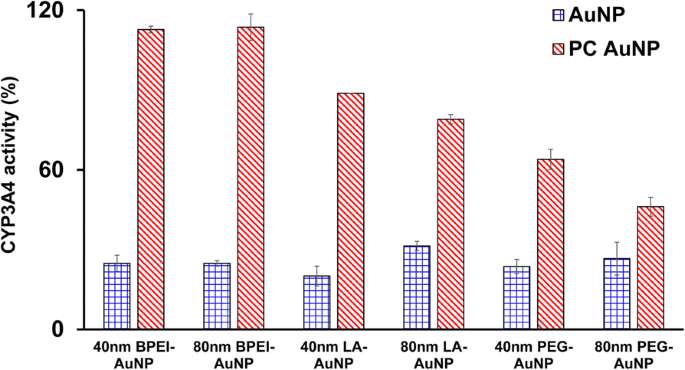
CYP3A4 활동 측정
CYP3A4 활성에 대한 40 및 80nm 베어 및 PC AuNP의 억제 효과를 특성화했습니다. 그림 5와 같이 LC50에서 40nm BPEI-, LA-, PEG-AuNP와 80nm BPEI-AuNP 값은 크기와 표면 전하에 관계없이 대조군에 비해 20.1~31.4%의 상응하는 활성으로 C3A 세포에서 CYP3A4의 촉매 활성을 억제했습니다. 80nm LA- 및 PEG-AuNP의 무독성 농도도 활성을 억제했습니다(각각 31.4% 및 26.6%). 그러나 PC는 대조군과 비교하여 63 및 46% 활성을 나타내는 40 및 80 nm PEG-AuNP 외에 40 및 80 nm AuNP 유도 CYP3A4 억제를 광범위하게 개선했습니다. 이것은 음이온성 탄닌산-AuNP와 양이온성 40 및 80nm BPEI-AuNP가 CYP3A4의 촉매 활성을 실질적으로 억제한다는 인간 간 조직 및 간세포에 대한 시험관 내 연구와 일치합니다[7, 25]. 대조적으로, 양이온성 PEI-AuNP와 중성 폴리비닐피롤리돈-AuNP는 HepG2 세포에서 CYP1A2, CYP2C9, CYP3A4, 쥐 간 절편에서 각각 CYP2B와 CYP3A의 mRNA 발현을 유도했다[36, 37]. 최근 연구에 따르면 40 및 80nm 베어 및 PC BPEI-AuNP는 단백질의 구조적 변화를 통해 인간 간세포에서 CYP3A4 활성을 실질적으로 억제하거나 가역적 억제로서 기질 포켓을 차단한다고 보고합니다[7].
<그림><그림>
24시간 동안 PC의 부재 및 존재 하에 40 및 80nm BPEI-, LA- 및 PEG-AuNP에 노출된 C3A 세포에서 CYP3A4 활성에 대한 AuNP의 억제 효과. 값은 평균 ± S.D를 나타냅니다. (n =3). BPEI 분지형 폴리에틸렌이민, LA 리포산, PEG 폴리에틸렌 글리콜, PC 인간 혈장 단백질 코로나
40nm PEG-AuNP의 독성 경로 집중 유전자 발현 프로파일링
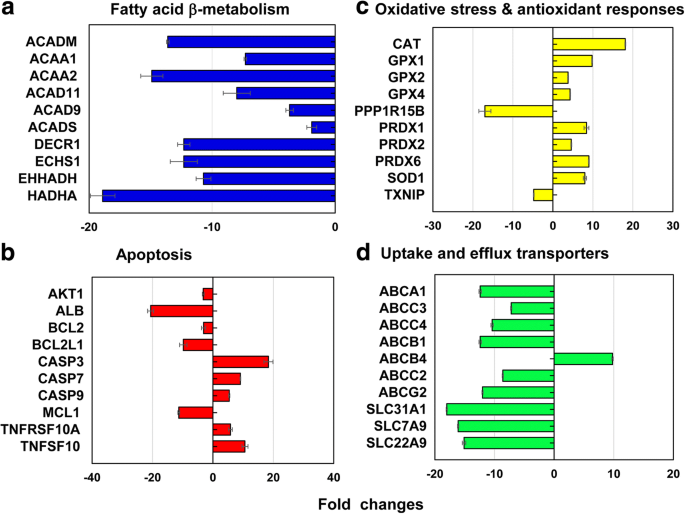
13개의 서로 다른 스트레스 및 독성 경로를 포괄하는 대표적인 유전자 중에서 총 212개의 유전자(↓186 및 ↑26 유전자)가 LC50에서 차등적으로 발현되었습니다. 40nm PEG-AuNP의 값(그림 6, 추가 파일 1:표 S2–S7). 전체 유전자(212개 유전자) 중 12.3%(26개 유전자, ↓26개, ↑0개 유전자)가 미토콘드리아 지방산 β-산화에 주로 관여했습니다. 세포자멸사 11.3%(24개 유전자, ↓18개, ↑6개 유전자); DNA 손상 및 복구 경로 11.3%(24개 유전자, ↓18, ↑6개 유전자); 그리고 열충격 반응의 경우 11.3%(24개 유전자, ↓22, ↑2).
<그림><그림>
a에 관여하는 대표적인 유전자 미토콘드리아 지방산 β-산화, b 아폽토시스, c 산화 스트레스 및 항산화 반응, d LC50에서 간 흡수 및 유출 수송체 40nm PEG-AuNP 값 모든 데이터는 p에서 배수 변화가 <− 2 및> 2였습니다. <0.05. 유전자 온톨로지 분석은 추가 파일 1에 나열되어 있습니다. 표 S2–S7
미토콘드리아 지방산 β-산화 경로에서 아실-CoA 생성에 관여하고 NADH 및 FADH2의 환원 당량에 관여하는 세 가지 다른 효소를 인코딩하는 유전자 주로 억압되었다; Acyl-CoA 탈수소효소의 ACAD11, ACAD9, ACADM 및 ACADS 유전자(2.0-13.6배); 케토아실-CoA 티올라제에서 ACAA1 및 ACAA2(7.3~14.9배); enoyl-CoA hydratase의 DECR1, ECHS1, EHHADH 및 HADHA(10.7~18.9배)(그림 6a, 추가 파일 1:표 S2). 미토콘드리아 지방산 β-산화는 아실-CoA 생성과 NADH 및 FADH의 당량 감소에 중요한 역할을 합니다2 , 이는 4가지 주요 효소(아실-CoA 탈수소효소, 에노일-CoA 수화효소, 히드록시아실-CoA 탈수소효소, 케토아실-CoA 티올라제[38, 39])와 관련이 있습니다. 또한, 전자 운반자, NADH 및 FADH2 , 트리카르복실산(TCA) 회로와 미토콘드리아 호흡 사슬에 관여하여 ATP 생성을 유발합니다. 현재 연구에서 40nm PEG는 세포 내 ATP 및 FADH 수준의 감소를 통해 ATP 유지의 손실인 미토콘드리아 기능 장애를 유도했습니다2 결과적으로 C3 세포에서 세포 독성을 정의합니다 (그림 2a). 유사한 현상이 40nm BPEI-AuNP에 노출된 인간 간세포, HUVEC 및 HPTC에서 보고되었으며, 이는 미토콘드리아 기능 장애가 표면 전하 및 세포 유형에 관계없이 AuNP 독성의 일반적인 메커니즘일 수 있음을 나타냅니다[7,8,9]. 최근 연구에 따르면 STAT3 인산화 억제제인 OPB-51602에 반응하여 불멸화된 전립선암 상피 세포와 폐암 상피 세포에서 미토콘드리아 기능 장애 관련 세포 독성이 관찰되었다고 보고되었습니다[40].
세포 사멸 경로에서 CASP3, CASP7, CASP9, TNFRSF10A, TNFRSF10B 및 TNFSF10의 6개 세포 사멸 촉진 유전자는 상향 조절된 반면, AKT1, ALB, BCL2, BCL2L1, MCL1 및 XIAP의 6개 항 세포 사멸 유전자는 하향 조절되었습니다(그림 1c). . 6b, Additional file 1:Table S3), which was correlated with dose-dependent cytotoxicity in C3A cells (Fig. 2a). In DNA damage and repair check point, genes of the checkpoint kinases (CHEK1/2), the DNA excision repair genes (ERCC1/2/3), and the DNA ligase IV (LIG4) were upregulated but other excision repair genes (ERCC5/6, XRCC1/5), the checkpoint kinase (CDKN1A), and protein kinases (PRKDC) genes were downregulated (2- to 19-fold). These results suggested that the 40 nm PEG-AuNP-induced interference with cell cycle and DNA repair system may correlate with an induction of cell death in C3A cells (Fig. 2a, Additional file 1:Table S3). Genes encoding two different heat shock proteins (HSP) (A1A and A1B) were upregulated (10.2- to 14.2-fold) but HSP40 subfamily A, B, and C; HSP90 member 1; and HSP60 were downregulated (2- to 16-fold) (Additional file 1:Table S4).
In oxidative stress and antioxidant response, the 40 nm PEG-AuNP at LC50 value induced antioxidants genes and suppressed pro-oxidants, which was associated with a decrease in ROS/RNS generation being antioxidant itself (Fig. 4b). In antioxidant genes, glutathione peroxidase (GPX) 1, GPX2, GPX4, PRDX1, PRDX2, PRDX6, superoxide dismutase (SOD) 1, and CAT were induced (3.8- to 18.1-fold). In pro-oxidant genes, TXNIP and PPP1R15B were suppressed (4.8- and 17-fold, respectively) (Fig. 6c, Additional file 1:Table S5). This is consistent with a previous study that AuNP displayed oxidative stress-induced cytotoxicity in HepG2 and human hepatocytes, irrespective of size [7, 19].
In phase I metabolism, CYP3A4 and ESD genes were extensively suppressed (7-fold and 12-fold, respectively). Especially, inhibitory effect of 40 nm PEG-AuNP on CYP3A4 expression was correlated with a decrease in CYP3A4 activity (Fig. 5). Recent studies reported that the 40 nm BPEI-AuNP inhibited gene expression of CYP1A2, CYP2C9, and CYP3A4 in human hepatocytes; ESD in HUVEC; and CYP1A1 in HPTC [7,8,9]. Epidemiology study demonstrated that CYP enzymes in liver tissue of HCC patient were substantially inhibited by the tumorigenic process at the molecular and the functional level [41].
Drug Uptake and Efflux Transporter Gene Expression Profiling
The development of multidrug resistance (MDR) by tumor cells is one of the main causes of cancer treatment failures [42, 43]. Integral membrane transporters-mediated decrease in drug uptake and increase in drug efflux including P-glycoprotein (P-gp) and breast cancer resistance protein (BCRP) is one of the major mechanisms of MDR.
Differential gene expression of drug efflux and uptake transporters in C3A cells exposed to 40 nm PEG-AuNP showed that a total of 14 genes of ABC transporters (↓12 and ↑2 genes) and a total of 21 genes of SLC transporters (↓21 and ↑0 genes) were substantially modulated at LC50 value (Figs. 6d and 7, Additional file 1:Table S7). In drug efflux transporters of ABC family, genes of multidrug resistance-associated protein (MRP3/ABCC3), MRP4 (ABCC4), and cholesterol efflux regulated protein (CERP/ABCA1) in basolateral membrane were downregulated (7.2- to 10.4-fold). The genes encoding P-gp (ABCB1), MRP2 (ABCC2), BCRP (ABCG2), and sterolin 2 (ABCG8) in canalicular efflux transporters were also suppressed (8.6- to 13.8-fold). In contrast, multidrug resistance (MDR4/ABCB4) in canalicular membrane and mitochondria ABC transporter (MTABC3/ABCB6) in the outer mitochondrial membrane were highly upregulated (9.8-fold and 5.8-fold, respectively). In drug uptake transporters, genes of copper transporter protein (CTR1/SLC31A1) and to a lesser degree organic anion transporting (OAT7/SLC22A9) were also inhibited (18- fold and 15-fold, respectively). These results support a recent study that the 40 nm BPEI-AuNP downregulate MDR3 in human hepatocytes but upregulates MRP3 in HUVEC indicating surface charge- and cell type-dependent interaction between AuNP and efflux transporters [7, 8]. Epidemiology study exhibited that a high expression of BCRP and a low expression of OCT3 occurred in HCC tumor, which was closely associated with the tumor progression and its size [44]. A previous study exhibited that P-gp inhibitor, verapamil enhanced cytotoxicity of glutathione-AuNP conjugated with doxorubicin in feline fibrosarcoma cell lines by increasing intracellular drug concentration [45]. The current study emphasizes that the mechanisms-derived information on the 40 nm PEG-AuNP identified a separate but still complementary action on mitochondrial fatty acid β-oxidation, TCA cycle and respiratory chain, drug efflux and uptake transporters, as well as CYP3A4 activity in C3A cells (Fig. 7). To the end, this will highlight AuNP interaction with key biological processes and its underlying molecular mechanism in HCC, which may be further implicated in the development of more effective therapeutic target in HCC treatment.
A schematic representation of the basic mechanisms of action of 40 nm PEG-AuNP in HCC treatment. Green bars (an inhibition) and pink triangles (an induction) indicate the 40 nm PEG-AuNP-modified biological markers and pathways. Gene ontology analysis is listed in Additional file 1:Tables S2–S7
To validate gene expression analysis from RT
2
array, the nine genes were selected for real-time PCR. In Additional file 1:Table S1, all nine genes were modulated at LC50 of the 40 nm PEG-AuNP. These transcriptional changes were consistent with those in gene expression analysis with PCR arrays (Fig. 6, Additional file 1:Tables S2–S7).
Conclusions
We have presented that cationic BPEI-, anionic LA-, or neutral PEG-AuNP interaction with human plasma protein corona (PC) caused the changes in DH , PDI, and z-potential of AuNP and further influenced cellular responses in C3A cells. All bare (no PC) 40 and 80 nm AuNP were cytotoxic to C3A cells besides the 80 nm LA-AuNP but PC completely ameliorated their cytotoxicities besides the 80 nm BPEI-AuNP. The 40 nm bare BPEI-AuNP showed the highest cellular uptake followed by the 40 nm PEG-AuNP and then the 40 nm LA-AuNP, whereas PC suppressed AuNP uptake besides the 80 nm PEG-AuNP. The 40 nm BPEI-AuNP caused biphasic responses of oxidative stress (pro- and antioxidant) in C3A cells, whereas the 40 nm PEG-AuNP was antioxidant. CYP3A4 activity was extensively suppressed by all bare AuNP, irrespective of size and surface charges, whereas PC substantially ameliorated its inhibitory effect on enzyme activity besides the 40 and 80 nm PEG-AuNP. Differentially expressed genes at LC50 value of 40 nm PEG-AuNP were mainly involved in mitochondrial fatty acid β-oxidation and to a lesser degree hepatic efflux/uptake transporters. The 40 nm PEG-AuNP inhibited three main enzymes in β-oxidation (acyl-CoA dehydrogenase, enoyl-CoA hydratase, and ketoacyl-CoA thiolase), other enzymes in TCA cycle, and the mitochondrial respiratory chain for ATP production. The 40 nm PEG-AuNP increased the expression of pro-apoptotic genes and decreased anti-apoptotic genes at LC50 값. A high level of antioxidants and a low level of pro-oxidants genes were observed in C3A cells exposed to 40 nm PEG-AuNP. In addition, genes of drug efflux and uptake transporters located in both basolateral and canalicular membrane were substantially modulated.