인간 지방 유래 줄기 세포를 연골로 분화시키는 원자력 현미경 기반 나노스코피:나노구조 및 인테그린 β1 발현
초록
Integrin β1은 분화, 이동, 증식, 상처 복구, 조직 발달 및 기관 형성에 관여하는 것으로 알려져 있습니다. 인테그린 β1 리간드와 분화 29(CD29) 수용체 클러스터 간의 결합 확률을 분석하기 위해 원자간력현미경(AFM)을 사용하여 인간 지방유래 줄기세포(hADSc) 표면의 천연 인테그린 β1 결합 수용체를 검출했습니다. . 인테그린 β1 리간드-수용체 상호작용의 결합 확률은 2차원 세포 배양 수준에서 초기 연골 형성 분화 동안 hADSc에 대한 인테그린 β1 기능화된 팁에 의해 조사되었습니다. hADSc의 세포 형태 및 미세 구조는 AFM으로 측정되었으며, 이는 긴 방추 세포가 연골 형성 유도 동안 길이/너비 비율이 감소하고 거칠기가 증가된 다각형 세포가 되었음을 입증했습니다. 인테그린 β1 리간드와 CD29 수용체의 결합은 살아있는 hADSc에 대한 β1 기능화된 팁에 의해 검출되었습니다. 연골 유도 0, 6, 12일에 총 1200개의 곡선이 기록되었습니다. 평균 파단력은 각각 61.8 ± 22.2pN, 60 ± 20.2pN, 67.2 ± 22.0pN이었다. 파열 사건은 각각 19.58 ± 1.74%, 28.03 ± 2.05%, 33.4 ± 1.89%였으며, 이는 연골 형성 유도 동안 hADSc 표면의 수용체와 인테그린 β1 리간드 사이의 결합 확률이 증가함을 입증했습니다. Integrin β1과 β-catenin/SOX 신호 전달 경로는 연골 분화 동안 상관관계가 있었습니다. 이 조사의 결과는 AFM이 연골 형성 동안 hADSc에 결합하는 인테그린 β1 리간드-CD29 수용체의 변화에 대한 운동 및 시각적 통찰력을 제공한다는 것을 의미합니다. 세포 형태, 막 미세구조 및 리간드-막횡단 수용체 결합 확률의 변화는 연골 분화 과정의 평가에 유용한 마커인 것으로 입증되었습니다.
<섹션 데이터-제목="배경">
배경
골관절염(OA)은 관절 연골의 점진적인 파괴를 특징으로 하는 퇴행성 골관절염과 함께 노인들에게 흔한 퇴행성 관절 질환입니다[1]. 연골은 혈관, 신경 또는 림프 조직 없이 고도로 조직화되어 있습니다[2]. 세포외기질(ECM)은 주로 콜라겐 II와 당단백질로 구성되어 있으며 연골 항상성에 매우 중요합니다. 연골은 혈관이기 때문에 자기 재생 능력이 제한적입니다. 골관절염 치료(외과적 및 비수술적)는 골관절염 환자의 증상, 특히 통증을 신속하게 완화할 수 있지만 관절 연골의 정상적인 구조와 기능을 회복할 수 없습니다[3]. 향후 치료에는 줄기세포를 이용한 조직 공학과 결손 및 퇴행성 관절 연골을 복구하기 위한 스캐폴드가 포함될 것입니다[4]. 간엽줄기세포는 성장인자의 조합에 따라 골형성, 지방형성, 연골형성, 근형성 잠재력을 갖는 다능성 기질세포이다[5]. 중간엽 줄기세포 분화 분석은 Wnt/β-카테닌, 포유류의 라파마이신 표적(mTOR), 포스포이노시티드 3-키나제(PI3K) 및 기타 경로가 분화에 중요한 역할을 하는 것으로 나타났습니다[6,7,8]. 그러나 연골 분화가 유도되는 기본 메커니즘은 아직 파악하기 어렵습니다. 이것은 세포외 신호가 세포내 신호전달 경로를 활성화시키는 메커니즘에 특히 해당됩니다. 연골 분화 동안 인테그린 β1이 변화하는 것을 발견했습니다. 따라서 우리는 인테그린 β1이 다양한 조직 분화 신호 전달 경로에 관여하기 때문에 인간 지방 유래 줄기 세포(hADSc) 연골 분화에서 중요한 역할을 할 수 있다고 가정했습니다. 이 조사에서는 Wnt/β-카테닌 신호 전달 경로에 초점을 맞추었습니다.
수많은 연구에서 세포와 세포외 환경 간의 상호작용이 막횡단 단백질, 특히 인테그린 계열 구성원에 의해 조절된다는 사실이 밝혀졌습니다[9]. 인테그린은 비공유 결합된 α 및 β 사슬의 이종이량체-막횡단 당단백질로 구성됩니다[10]. 이론적으로 64개의 알려진 인테그린이 있으며 그 중 24개만 발견되었습니다. 인테그린은 세포-세포 접착, ECM-세포 접착, 세포 신호 전달 및 액틴 세포골격의 구성에 중요한 역할을 합니다[11]. ECM은 조직 항상성에서 중요한 역할을 하며 ECM은 인테그린을 조절합니다. 인테그린은 세포 접착, 이동, 증식, 분화, 세포 사멸, 상처 복구, 조직 발달 및 기관 형성을 포함한 많은 기본 과정을 매개합니다. 중간엽 줄기세포 연골형성 분화 동안, 인테그린 β1의 발현은 SOX 신호전달 경로 및 콜라겐 II에 연결된다. 인테그린 β1 이합체는 연골 이종이량체 중에서 가장 두드러지고 다양한 α 이량체와 상호작용하는 것으로 알려져 있기 때문에 이 연구의 초점은 β1 이합체에 있었습니다[12]. 분화 클러스터 29(CD29)는 거의 모든 세포 및 조직 유형에서 발현되는 매우 늦은 항원 수용체와 관련된 인테그린 β1 소단위입니다.
여기에서 원자간력현미경(AFM)을 사용하여 hADSc 연골 분화 동안의 변화를 측정할 수 있었습니다. 매우 고해상도 유형의 스캐닝 프로브 현미경 검사법으로서 AFM은 나노 규모의 유체에서 단일 세포의 형태와 세포막을 검출할 수 있는 새로운 기회를 제공했습니다. 한편, 원자간력현미경(AFM)과 결합된 단일 분자 힘 분광법(SMFS) 시스템은 살아있는 세포에 대한 리간드-수용체 결합을 측정하는 데 사용되었습니다. SMFS의 시스템은 세포막의 수용체 변화에 더 민감하고 결합력의 이미지가 시각화되었습니다. 이 작업에서 인테그린 β1 리간드-수용체 결합은 인테그린 β1 기능화된 AFM 팁에 의해 조사되었습니다. AFM을 적용하면 연골 분화가 hADSc 세포 모양을 변화시키고 세포 거칠기를 증가시키는 것으로 밝혀졌습니다. 이 응용 프로그램은 인테그린 β1 리간드-수용체 상호 작용 및 세포 표면 미세 구조 변경을 직접 측정하여 연골 분화를 평가하는 방법을 제공하여 시각화된 방식으로 세포 표면 조사 및 스크리닝을 개선했습니다. 연골 형성 분화는 세포 내 세포 골격 상호 작용뿐만 아니라 막 구성 및 구조를 변경합니다. 세포 형태, 미세구조 및 리간드-막횡단 수용체 결합의 이러한 변화는 연골 형성 분화 메커니즘의 평가에 유용한 마커 역할을 합니다.
방법
세포 배양 및 시약
이 조사를 위해 이전에 설명한 대로 3명의 수술 환자(평균 연령 20세)에서 세포를 분리했습니다[13]. 모든 환자로부터 사전 동의를 받았습니다. 본 연구에 대한 윤리 승인은 지난대학교 제1부설병원(보충서식)에서 받았습니다. 세포는 10% 열-불활성화 소태아혈청(FBS, Life Technologies, CA, USA)이 보충된 저포도당 Dulbecco's Modified Eagle's Medium(DMEM, Life Technologies, CA, USA), 100 units/ ml 페니실린(Life Technologies, CA, USA), 100μg/ml 스트렙토마이신(Life Technologies, CA, USA), 0.11mg/ml 소듐 피루브산(Life Technologies, CA, USA) 및 L-글루타민(Life Technologies, CA, 미국). 세포는 5% CO2를 포함하는 가습 인큐베이터에서 37°C로 유지되었습니다. 3일마다 매체가 변경됩니다.
체외 분화
연골 형성 유도를 위해 4~8계대 hADSc를 높은 세포 밀도(2 × 105/10 ml)로 파종하고 1% FBS, 1% 인슐린-트랜스페린-셀레늄( ITS) + 보충제(Cyagen, 중국 광저우), 10ng/ml 변형 성장 인자-베타1(TGF-β1)(Peprotech, Rocky Hill, New Jersey, USA), 100ng/ml 인슐린 유사 성장 인자-1( IGF-1)(Peprotech, Peprotech, Rocky Hill, New Jersey, USA), 10-7 M 덱사메타손(Sigma, St. Louis, MO, USA) 및 50μg/ml 아스코르브산(Sigma, St. Louis, MO) , 미국). TGF-β1 및 IGF-1을 새로 추가하여 배지를 2일마다 교체했습니다. 연골 형성은 알시안 블루 및 톨루이딘 블루 염색으로 평가되었습니다.
골형성 및 지방형성 분화를 유도하기 위해 4-8계대 세포에 각각 2주 동안 골형성 및 지방형성 배지를 처리하였다. 골형성 배지는 10-7 M 덱사메타손(Sigma, St. Louis, MO, USA), 50μg/ml 아스코르브산(Sigma, St. Louis, MO, USA) 및 10mmol/l β-글리세롤이 보충된 DMEM으로 구성되었습니다. 포스페이트(Sigma, St. Louis, MO, USA). 골형성은 알리자린 레드 염색으로 평가했습니다.
지방 생성 배지는 0.5mmol/l 3-이소부틸-1-메틸크산틴(IBMX)(Sigma, St. Louis, MO, USA), 1μmol/l 하이드로코르티손(Sigma, St. Louis, MO, USA)이 보충된 DMEM으로 구성되었습니다. 0.1mmol/l 인도메타신(Sigma, St. Louis, MO, USA). Oil Red O 염색으로 지방 분화를 평가했습니다.
유세포분석을 통한 hADSc 표면 항원 식별
hADSC를 트립신으로 분해한 다음 DMEM으로 두 번 세척한 다음 2 × 10
7
의 세포 밀도에서 재현탁했습니다. 세포/ml. 세포 현탁액(50μl, 1 × 10
6
세포)를 1.5ml 에폭시 에폭사이드 튜브에 첨가한 다음 항-CD34, 항-CD44, 항-CD45, 항-CD73, 항-CD90, 항-CD106, 항-HLA-DR 및 항-CD105 항체와 함께 배양했습니다. 어둠 속에서 37°C에서 20분 항-CD34, 항-CD44 및 항-CD45는 CST(미국 매사추세츠주 비벌리)에서 입수했습니다. 다른 항체는 Abcam(Cambridge, MA, USA)에서 입수했습니다. 그런 다음, 세포 현탁액을 × 500g에서 원심분리했습니다. 5분 동안 처리한 후 상층액을 제거하고 200μl의 Stain Buffer에 세포를 재현탁합니다. 모든 단계는 유세포 분석에 의한 분석 전에 두 번 반복되었습니다.
면역블롯팅 분석(IB)
이전에 설명한 대로 면역블롯팅을 위해 세포를 수집했습니다[14]. 사용된 1차 항체는 Abcam(Cambridge, MA, USA)에서 입수한 항-β-카테닌(ab32572), 항-인테그린 β1(ab30394) 및 항-콜라겐 II(ab34712)였습니다. 항-β-액틴(8H10D10, 1:2000), 항-GSK-3β(27C10, 1:1000) 및 항-SOX(92G2, 1:1000)는 Cell Signaling Technology(CST, Beverly, MA, 미국). 2차 HRP 결합 항체(1:1000–1:3000)는 CST에서 구입했습니다.
면역형광
연골 분화를 위해 세포를 0, 6 및 12일 동안 처리하고 소화하고 24웰 플레이트(Costar353047, Corning, New York, USA)의 유리에서 24시간 동안 배양했습니다. 세포를 얼음처럼 차가운 인산염 완충액(PBS)으로 두 번 세척하고 실온에서 15분 동안 4% 파라포름알데히드로 고정했습니다. 차단 후, 세포를 인테그린 β1과 반응성인 1차 항체와 함께 1시간 동안 배양한 다음, Alexa Fluor 488 표지된 항-마우스 IgG(H + L)(CST #4408, MA, USA)와 함께 암실에서 1시간 동안 배양했습니다. ), 4',6-디아미디노-2-페닐인돌(DAPI, Sigma, MO, USA). 팔로이딘 염색의 경우 차단 후 세포를 0.2% Triton X-100으로 30분 동안 투과화한 다음 DAPI 및 팔로이딘-Alexa Flour 573(Life Technologies, CA, USA)과 함께 1시간 동안 배양했습니다. 세 번 세척한 후, 연골 분화 동안 인테그린 β1의 세포내 위치와 사상액 액틴(F-액틴)의 변화를 레이저 스캔 공초점 현미경(ZEISS, LSM 700, Oberkochen, Germany)으로 평가했습니다.
AFM 팁 준비
스프링 상수(0.06N/m)를 갖는 Si3N4 팁(DNP-10, Bruker Corp)은 다음과 같이 항-CD29 항체에 의해 화학적으로 변형되었다[15]. 팁은 아세톤, 자외선 및 피라냐 용액(H2 SO4 :H2 O2 =3:1, v /v ) 다양한 시간(5분, 30분, 10분) 정제수로 철저히 헹군 후 에탄올 중 1% 3-APTES(Sigma, St. Louis, MO, USA) 용액과 함께 30분 동안 배양하여 팁을 형성했습니다. 팁을 초순수로 3회 세척하고 2.5% 글루타르알데히드(Sigma, St. Louis, MO, USA) 용액으로 1시간 동안 처리했습니다. 불필요한 글루타르알데히드를 물로 3회 세척하였다. 마지막으로 팁을 항인테그린 β1 용액(1mg/ml)에 삽입하고 4°C에서 밤새 배양했습니다. 변형된 프로브는 실험 전에 PBS로 세척했습니다.
AFM 측정
AFM(Bioscope Catalyst, Bruker, USA)은 연골 분화 동안 hADSc 형태와 미세구조 변화를 조사하는 데 사용되었습니다. AFM 팁의 정확한 힘 상수는 PBS에서 측정되었습니다. 형태 및 미세구조를 평가하기 위해 세포를 PBS로 여러 번 세척했습니다. 그런 다음 4% 파라포름알데히드 용액을 3.5cm
2
15분 동안 배양 접시. 세포를 PBS로 세척한 후, 세포를 사용할 때까지 4°C에서 PBS에 보관했습니다. 팁의 스프링 상수는 접촉 모드에서 4.2~5.8N/m 범위였습니다. hADSc의 형태 및 미세 구조 이미지는 AFM에 의해 실온에서 PBS에서 촬영되었습니다. hADSc의 핵을 둘러싼 미세구조 이미지는 접촉 모드에서 획득되었습니다. 나노스코프 분석 소프트웨어를 사용하여 (0, 6, 12일차) 그룹에서 최소 15개의 서로 다른 세포에 대해 15개 이상의 서로 다른 10 × 10μm 이미지에 대한 세포 표면 미세구조를 평가했습니다. 인테그린 β1으로 변형된 AFM 팁과 살아있는 hADSc의 CD29 수용체 사이의 결합력은 다양한 연골형성 기간(0, 6, 12일) 동안 분석되었습니다. 결합력은 AFM 시스템(Bioscope Catalyst, Bruker, USA)의 접근-후퇴 모드에서 측정되었다. 인테그린 β1-살아 있는 세포 분리 사건을 연구하기 위해 인테그린 β1 항체 수정 팁을 500nm/s의 접근-후퇴 속도에서 사용했습니다. 기능화된 팁의 힘 상수는 0.058 ± 0.006N/m였습니다. 세포에 대한 역치 힘은 800pN이었습니다. 힘 측정 실험 전에 항-인테그린 β1 항체(100μg/ml)를 30분 동안 세포에 첨가했습니다. 인테그린 β1 차단 및 베어 프로브도 인테그린 β1 항체 변형 팁과 세포 사이의 비특이적 파열력을 감지하기 위한 대조군으로 사용되었습니다. 인테그린 β1 리간드-수용체 결합 확률의 정량화를 위해 인테그린 β1 항체-기능화된 프로브에 의해 특이적 상호작용력 곡선을 측정하였다. 400개 이상의 힘 곡선이 최소 3개의 독립적인 실험에서 요약된 결과와 함께 단일 실험에서 측정되었습니다. 따라서 각 비교 실험에서 약 1200개의 원래 힘-거리 곡선이 기기의 나노스코프 분석 소프트웨어를 사용하여 30-40개의 서로 다른 셀에서 획득되었습니다. 최소 3번의 독립적인 실험에서 힘 값을 평균화하여 세포 표면에서 인테그린 β1 리간드와 CD29 수용체 사이의 상호 작용력에 대한 연골 형성 유도 효과를 결정했습니다.
역전사 및 Real-time PCR
TRIzol® Plus RNA Purification Kits(Life Technologies, CA, USA)를 사용하고 1μg의 RNA를 약간 수정하여 제조업체의 프로토콜에 따라 고용량 cDNA 역전사 키트(Invitrogen)를 사용하여 cDNA로 역전사했습니다. 인테그린 β1 및 GAPDH는 유전자 특이적 프라이머와 함께 qRT-PCR을 사용하여 정량화되었습니다:5'-TGGAGGAAATGGTGTTTGC-3'(인테그린 β1-센스) 및 5'-CGTTGCTGGCTTCACAAGTA-3'(인테그린 β1-안티센스); 5'-CTGACTTCAACAGCGACACC-3'(GAPDH-센스) 및 5'-CCCTTGTTGCTGTAGCCAAAT-3'(GAPDH-안티센스). Real-Time PCR을 위해 Fast SYBR@GREEN Master Mix(Life Technologies, CA, USA)를 사용하여 Step One Real-Time PCR(Applied Biosystems)을 수행했습니다. 표적 유전자 발현을 내부 표준으로서 GAPDH로 정규화하고 비교 2-ΔΔCT 방법을 사용하여 계산하였다. 각 분석은 세 번 수행되었습니다.
통계 분석
모든 실험은 평균 ± 표준편차(SD)로 표현된 데이터로 최소 3번 수행되었습니다. 두 그룹 간의 비교는 t에 의해 수행되었습니다. 테스트. 그룹 평균 간의 유의한 차이는 일원 ANOVA 분석에 의해 결정된 후 Bonferroni 및 Tamhane의 T2 테스트(등분산이 가정되지 않음)에 의해 결정되었습니다. p의 값 <0.05는 통계적으로 유의한 것으로 간주되었습니다.
결과 및 토론
hADSc 평가
간엽줄기세포는 골형성, 지방형성, 연골형성 및 근형성 잠재력을 갖는 다능성 기질 세포이다. hADSc, 세포 표면 CD 마커 및 분화 능력을 식별하는 두 가지 주요 수단이 있습니다[16]. 추가 파일 1:그림 S1 및 추가 파일 2:그림 S2에서 볼 수 있듯이 파생된 세포는 hADSc였습니다. 그런 다음, 계대 3 hADSc의 세포 증식은 MTT 분석에 의해 결정되었습니다(추가 파일 3:그림 S3).
hADSc 연골형성 동안 유도된 형태 및 표면 미세구조 변화
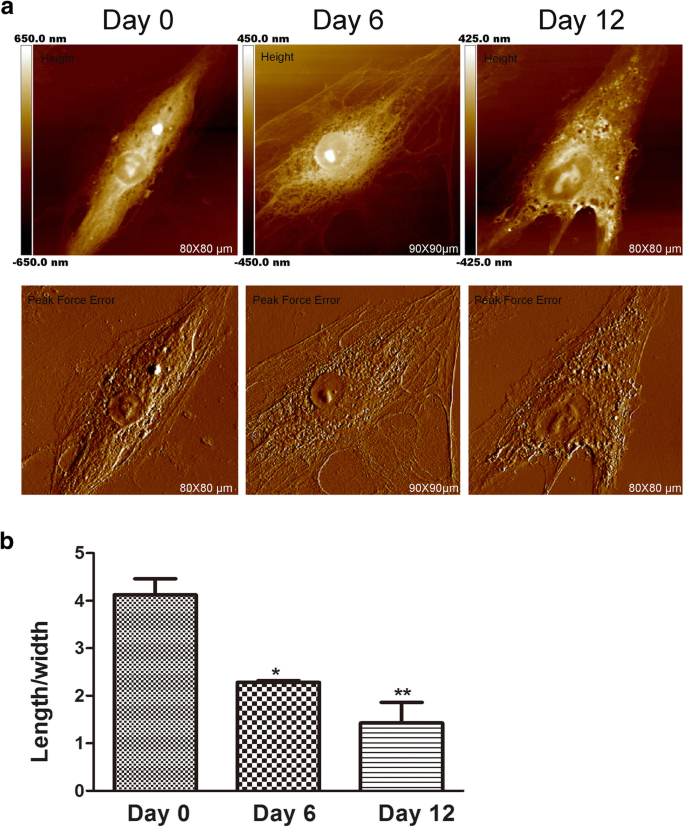
AFM은 항상 나노 규모에서 세포 형태와 미세 구조를 감지하는 데 사용됩니다[17]. 세포의 모양은 전문화된 세포 기능 및 조직 구성과 관련이 있습니다. 일부 암 연구에서 AFM은 약물 효과 평가를 위한 형태학적 변화를 분석하기 위한 고영상 기술로 사용될 수 있습니다. 또한 연골유도 과정에서 중간엽 줄기세포의 형태가 변한다[18]. 세포 모양의 변화가 분화에 필요한 것으로 보이지만 세포 형태가 중간엽 줄기 세포 분화의 초기 발달 단계에 영향을 미치는지 여부에 대해서는 알려진 바가 거의 없습니다. 따라서, hADSc 연골형성 동안의 형태 및 막 미세구조 변화는 AFM에 의해 평가되었는데, 이러한 변화는 중요하고[19], 세포의 기능에 직접 영향을 미칠 수 있기 때문이다[20]. hADSc의 표면 형태와 초미세 구조는 서로 다른 기간 동안 연골 분화 동안 조사되었습니다(그림 1 및 그림 2). 형태와 표면 미세구조는 각 비교군에서 분명히 달랐다. 0일째에, 세포는 비교적 매끄러운 표면을 갖는 긴 방추형을 가졌다. 세포막 구조는 균질했습니다. 연골 형성 유도 후, 6일과 12일에 상당한 세포 형태 변화가 관찰되었습니다. 대부분의 세포는 연골 분화 과정에서 평균 세포 길이/폭 비율이 감소하면서 점차 다각형 모양(그림 1a)으로 축소되었습니다(그림 1b). 수많은 연구에서 세포 형태의 변화가 세포의 세포골격과 일치함을 보여줍니다[21]. 우리는 또한 연골 분화 동안 세포 골격의 변화를 발견했는데, 이는 후자의 결과에서 설명되었습니다.
<사진>
연골 형성 중 hADSc 형태의 특성. 아 전체 hADSc의 형태학적 이미지는 연골 분화 0, 6, 12일에 얻었습니다. 이미지는 Nanoscope의 Height 및 Peak Force Error Image Model에 의해 분석되었습니다. ㄴ 세포의 평균 길이/폭 비율은 연골 분화 처리 후 0, 6 및 12일에 측정되었습니다. *p <0.05, **p <0.01
<그림>
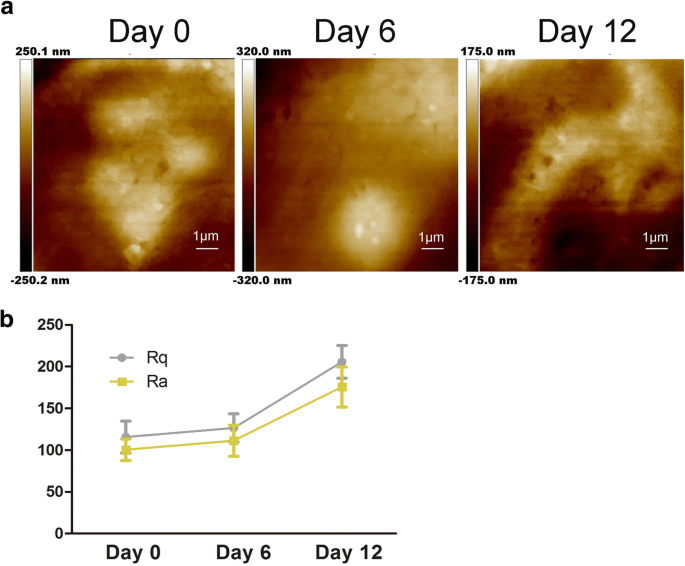
연골 분화 중 hADsc 막 미세 구조의 특성. 아 0, 6, 12일 동안 연골 분화 후 세포막 미세구조의 변화를 평가했습니다. ㄴ 세포의 표면 거칠기 매개변수 Ra 및 Rq는 0, 6 및 12일 동안 hADSc의 연골 형성 유도 동안 측정되었습니다.
그림 2a에서 볼 수 있듯이 세포막 미세구조도 변경되었습니다. 입자가 커지고 이질적이었습니다. 이전 연구에서는 Ra와 Rq가 다르게 처리된 세포막의 변화를 평가하기 위한 거칠기 값의 제작자임을 보여주었습니다[22]. Rq는 평균 제곱근 거칠기에 관한 것입니다. \( \mathrm{Rq}=\sqrt{\frac{\sum_{t-1}^N{\left( Zn-\overline{Z}\right)}^2 {N-1}} \); \( \mathrm{Rq}=\sqrt{\frac{\sum_{\mathrm{t}-1}^{\mathrm{N}}{\left(\mathrm{Zn}-\overline{\mathrm{Z }}\right)}^2}{\mathrm{N}-1}}; \) Ra는 평균 거칠기, \( \mathrm{Ra}=\frac{1}{N}{\sum}_{ t-1}^N1\mid Zi-\overline{Z}\mid\). 거칠기를 얻기 위해 스캔 크기는 10μm × 10μm입니다. 그림 2b에서 볼 수 있듯이, hADSc의 연골 형성 동안 두 개의 서로 다른 영역의 Ra와 Rq가 모두 증가했습니다. 0일째에 세포의 Ra 및 Rq 값은 낮아서 매끄러운 표면을 나타냅니다(그림 2b). Ra 및 Rq의 값은 연골 분화와 함께 동시에 증가하여 세포 표면에서 더 큰 이질성과 거칠기를 보여줍니다(그림 2a). 관찰된 변화에 기초하여 연골 분화는 세포 형태 및 세포 높이/너비 비율 변화를 초래했습니다(그림 1a, b). ECM이 인테그린을 조절하여 세포 접착을 조절할 수 있다는 연구 결과가 있습니다[11]. 따라서 증가된 거칠기 값은 연골 형성 동안 ECM과 세포막의 미세 구조의 변화를 시사합니다. 이 데이터는 세포 형태, ECM 및 세포막 구조에 영향을 미치는 연골 분화를 보여줍니다.
hADSc의 연골 형성 유도 중 세포골격 변화
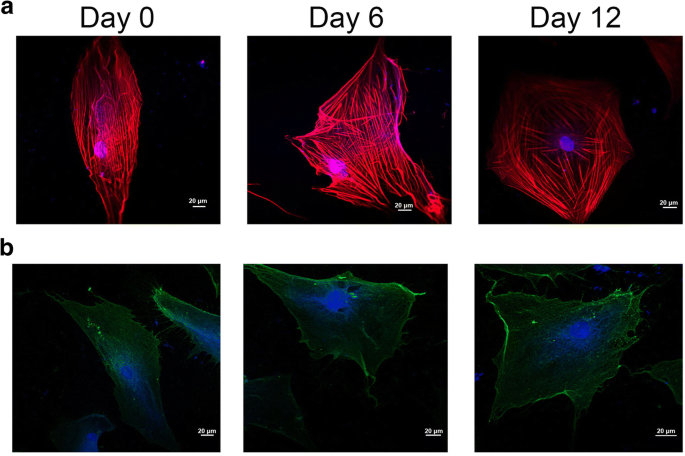
줄기세포 분화 과정에서 세포의 형태와 막의 구조적 변화는 세포의 세포골격과 관련되어 계통특이적인 세포적 특성이 발달하게 된다[21]. 도 3a에 도시된 바와 같이, 적색 및 청색 형광 신호는 각각 F-액틴 및 DAPI를 나타낸다. 세포 세포 골격은 그림 3a에서 연골 형성 유도 동안 크게 변화했습니다. 한편, 세포골격의 미세섬유는 0일째 군에서 장세포축을 따라 진행한 반면, hADSc를 12일 동안 연골 분화 처리했을 때 세포골격 미세섬유는 방사상 배열로 퍼졌다. 한편, 세포 미세섬유의 분포는 0일째 그룹에서 균질한 분포를 보였으나 12일 동안 연골 분화를 처리한 hADSc 주변부에 미세섬유가 주로 분포하였다.
<그림>
연골 분화 hADSc에서 세포 골격의 조직 및 인테그린 β1의 위치. 아 공초점 현미경으로 hADSc의 연골형성 동안 세포골격의 변화가 검출되었다. ㄴ 인테그린 β1의 위치는 연골 분화 동안 공초점 현미경으로 측정하였다. 세포골격과 핵은 각각 F-액틴과 DAPI로 염색되었다. 적색 및 청색 형광 신호는 각각 F-액틴 및 DAPI를 나타냅니다.
연쇄성 분화가 hADSc의 수용체에 대한 인테그린 β1의 결합 가능성을 변경
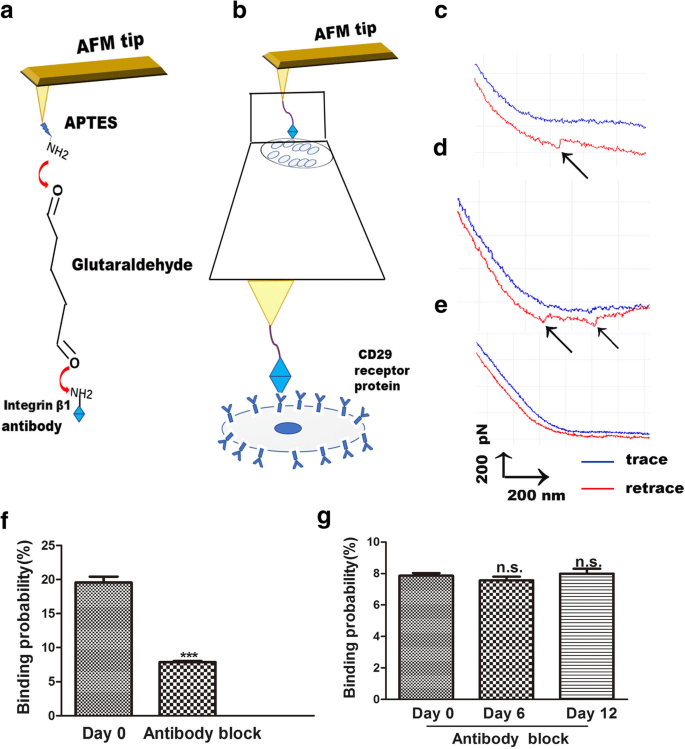
AFM은 또한 리간드와 그 수용체 사이의 결합력 연구에 유용한 도구로, 세포 표면의 막-수용체 신호 전달을 명확하게 합니다[23]. AFM에 의해 인테그린 β1과 그 수용체 사이의 변화는 시각적이고 간단하며 구체적인 방식으로 측정됩니다. 살아있는 세포에 대한 인테그린 β1 리간드-수용체의 상호작용은 세포막에 대한 결합 과정을 탐구하는 방법입니다. AFM 팁 기능화 절차는 APTES와 글루타르알데히드의 연결을 통해 인테그린 β1을 AFM 팁에 연결하는 것입니다. 이 팁은 세포 표면의 CD29 수용체에 대한 인테그린 β1의 결합을 감지하는 데 사용되었습니다(그림 4a). 단일 분자 힘 분광법(SMFS)을 사용하여 개별 살아있는 hADSc의 국소 영역 내에서 항인테그린 β1 살아있는 세포 분리력 분포를 평가했습니다(그림 4b). 대표적인 힘 곡선은 그림 4c, d에 나와 있으며, 이는 단일 분자 곡선(그림 4c)과 두 쌍의 파열 피크 곡선(그림 4d)을 나타냅니다. 얻어진 힘 곡선의 특이성을 확인하기 위해 차단 실험 및 베어 AFM 팁 실험을 수행했습니다. 노출된 원자현미경 팁은 특정 힘 피크를 감지하지 못했습니다(그림 4e). Bare AFM 실험은 hADSc 표면에서 integrin β1 리간드-수용체 상호작용의 비특이적 결합 확률이 1% 미만임을 보여주었다. 차단 실험을 위해 항인테그린 β1 항체를 세포와 함께 30분 동안 배양한 다음 인테그린 β1 기능화된 팁을 사용하여 힘 곡선을 기록했습니다. 차단 항체는 힘 곡선을 90%까지 감소시켰습니다(그림 4f). 항인테그린 β1 항체 처리 후 3군 간에 세포 표면에 대한 인테그린 β1 리간드-수용체의 결합 확률에는 차이가 없었다(그림 4g). 이러한 결과는 항체 변형 원자현미경 팁이 힘을 감지하는 데 매우 유용했으며 인테그린 β1 기능화된 원자현미경 팁이 특이적임을 보여줍니다.
<그림>
살아있는 hADSc에 대한 인테그린 β1 기능화된 원자현미경 팁을 사용한 원자현미경 힘 측정. 아 AFM 팁에 인테그린 β1을 고정하는 데 사용되는 전략의 도식적 표현. ㄴ 인테그린 β1 기능화된 AFM 팁과 살아있는 hADSc 사이에서 측정된 단일 분자 힘의 도식적 표현. c, d hADSc 및 e에 대한 인테그린 β1 수정 AFM 팁으로 얻은 대표적인 힘 곡선 인테그린 β1 모노클로날 항체 용액으로 시스템을 차단한 후. 에 0일에 인테그린 β1 항체에 의한 차단 전후의 hADSc에 대한 인테그린 β1 기능화된 팁의 결합 확률. g 0, 6, 12일에 인테그린 β1 항체로 차단한 후 hADSc에 대한 CD29 기능화된 팁의 결합 확률. ***p <0.001, n.s.. 큰 차이 없음
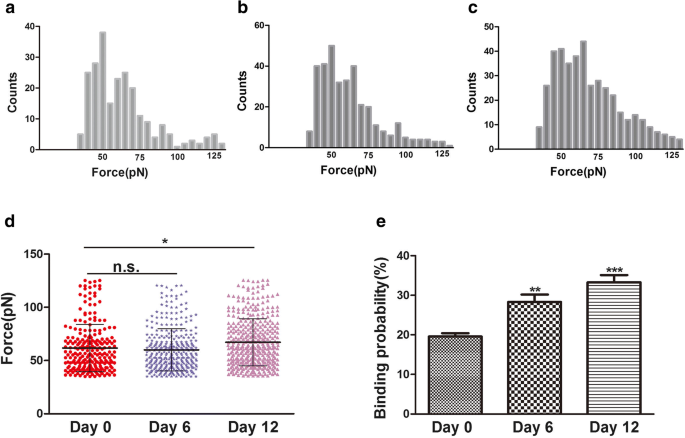
결합력(파열력)은 리간드와 그 수용체 사이의 상호작용력이다[24]. 원형질막의 형태 및 표면 미세구조의 변화는 분화, 세포자멸사 및 세포 이동과 같은 세포 생물학의 많은 과정과 관련이 있습니다. 분화 동안 세포골격의 변화는 인테그린 변화, 특히 인테그린 β1과 관련이 있는 것으로 생각됩니다. Integrin β1(CD29)은 ECM에 대한 세포 접착 및 세포-세포 접착에 매우 중요합니다. 또한 세포 내 단백질과 상호 작용하여 액틴 세포 골격과 관련된 신호 분자를 자극할 수 있습니다[25]. 이 연구에서 공초점 레이저 주사 현미경(CLSM)과 원자현미경(AFM)에 의해 hADSc 연골 형성 동안 세포골격 및 세포 형태 변화가 관찰되었습니다. 연골 분화 동안 세포골격, 형태 및 표면 미세구조의 변화는 세포 상태의 새롭고 신뢰할 수 있는 지표가 될 수 있습니다. CD29 수용체인 Integrin β1은 면역형광법에 의해 판단되는 바와 같이 세포 표면에 분포되어 있습니다(그림 3b). hADSc 연골 형성 동안 인테그린 β1 리간드-수용체 복합체의 결합 강도와 안정성을 분화 0, 6, 12일에 평가했습니다. 총 1200개의 곡선이 매일 기록되었으며 평균 파열력은 각각 61.8 ± 22.2pN, 60 ± 20.2pN 및 67.2 ± 22.0pN입니다(그림 5a–c). 힘 크기의 분포는 힘 평균 +로 분석되었습니다. SD(그림 5d). 0일과 6일 사이에 힘의 평균에는 큰 차이가 없었습니다. 0일과 12일 사이에는 힘 평균에 차이가 있었습니다. 결합력의 크기는 12일에 증가했습니다. 한편, 파열 사건은 0, 6, 12일 일에 각각 19.58 ± 1.74%, 28.03 ± 2.05%, 33.4 ± 1.89%였다(그림 5e). 증가된 결합 확률은 또한 인테그린 β1(CD29)이 연골 분화에서 중요한 역할을 했으며 신호 전달 경로를 통해 연골 분화에 대한 정보를 제공할 수 있음을 나타냅니다. 따라서 연골 분화 동안 증가된 인테그린 β1 나노도메인은 살아있는 hADSc에 대한 CD29 리간드-수용체의 결합 강도에 근본적으로 영향을 미칠 수 있습니다. 원형질막의 형태 및 표면 미세구조의 변화는 세포에 대한 인테그린 β1 리간드-수용체 복합체의 인테그린 β1 단백질 구조, 형태, 결합 강도 및 안정성의 변화를 동반하였다. 요약하면, 인테그린 β1은 hADSc 연골 분화에 필수적인 역할을 합니다.
<그림>
인테그린 β1 기능화된 AFM 팁에 의해 살아있는 hADSc의 표면에서 측정된 결합력 및 결합 확률. a–c 0, 6, 12일 동안 hADSc 연골 분화 동안 얻은 인테그린 β1 항체-수용체 결합력의 히스토그램. d 인테그린 β1 수용체에 대한 결합력은 hADSc의 연골 분화 0, 6, 12일에 얻어졌습니다. 이 인테그린 β1-수용체의 결합 확률은 0, 6 및 12일 동안 hADSc의 연골 분화 동안 검출되었습니다. *p <0.05, **p <0.01, ***p <0.001, n.s. 큰 차이 없음
hADSc 연골 분화 동안 인테그린 β1의 상향 조절
수많은 연구에서 인테그린 계열 구성원이 세포 분화에 중요한 역할을 한다는 것이 밝혀졌습니다. 또한, 인테그린은 세포외 환경과 세포 사이의 상호작용을 조절하여 연결된 단백질을 통한 신호 전달 경로를 제어할 수 있습니다[26]. 이전 연구에서는 결합 확률이 세포 표면의 막횡단 단백질(수용체)의 밀도와 형태에 영향을 받을 수 있음을 보여주었습니다[27]. 인테그린 형태는 리간드에 대한 친화도가 낮은 폐쇄형 헤드피스 또는 리간드에 대한 친화도가 높은 개방형 헤드피스일 수 있습니다[28, 29]. 인테그린 β1의 발현은 연골 세포의 특징인 콜라겐 II 발현의 증가와 함께 전사 및 번역 수준 모두에서 상향 조절되었습니다(그림 6a, b). 이와 같이 상향 조절된 인테그린 β1 발현은 형태에 관계없이 결합 확률 증가와 일치했습니다.
The role of integrin β1 and β-catenin/SOX pathway in regulating hADSc chondrogenic differentiation. 아 Protein integrin β1 was up-regulated during chondrogenesis of hADSc as assessed by western blotting. Cartilage differentiation up-regulated collagen II expression at different days. ㄴ The mRNA of integrin β1 was up-regulated during chondrogenic differentiation of hADSc. ㄷ Measurement of proteins associated with the β-catenin/SOX pathway during chondrogenic differentiation of hADSc for 0, 6, and 12 days. *p <0.05, **p <0.01
The Role of Integrin β1 in Chondrogenic Differentiation Regulated by the β-catenin/SOX Signaling Pathway
Previous studies have shown Wnt/β-catenin, PI3K, and mTOR signaling pathways to be related to integrin β1 [30,31,32]. Each is important in mesenchymal stem cell differentiation. Likewise, studies have demonstrated SOX and collagen II to be regulated by integrin β1 during chondrogenesis of hADSc. SOX is a hallmark component of the Wnt/β-catenin signaling pathway. Hence, we hypothesized that chondrogenic differentiation was regulated by the β-catenin/SOX pathway via integrin β1. SOX, GSK-3β, β-catenin, and integrin β1 were all increased during chondrogenesis of hADSc (Fig. 6c), with integrin β1 inducing cell signaling. These data demonstrate chondrogenic differentiation to be regulated by the β-catenin/SOX pathway via integrin β1.
Prospective and Limitations
In this work, changes in cellular morphology, the structure of the membrane, and the binding probability of integrin β1 ligand–receptors were demonstrated to be useful image markers to evaluate the chondrogenic differentiation process. This is a new method for evaluation of morphology, membrane ultrastructure, and changes in transmembrane proteins during chondrogenic differentiation. There are limitations to this study. Although increased binding probability was related to the high expression of integrin β1, the conformation of integrin β1 during chondrogenesis was not investigated. Further work is necessary to determine the conformation of integrin β1 during chondrogenic differentiation. Integrin β1 was demonstrated to participate in the β-catenin/SOX signaling pathway during chondrogenesis of hADSc. However, the relationship between integrin β1 and β-catenin/SOX signaling pathway is still not fully established. Further work is necessary to identify the exact role of integrin β1 in this pathway.
결론
In the present work, a novel method (AFM) was employed to evaluate chondrogenic induction in hADSc. Cell surface ultrastructural changes were assessed by AFM imaging. AFM was used to investigate the binding force and binding probability between integrin β1 ligand and its receptors on the surface of hADSc by integrin β1-functionalized AFM tips. Based on AFM data, during chondrogenesis, cell morphology was changed from an elongated spindle shape to a polygonal shape with increased cell roughness. By use of integrin β1-functionalized AFM tips, the binding probability and force magnitude of integrin β1 ligand–receptor on the surface of hADSc were found to increase during chondrogenic induction. By immunoblot, integrin β1 was demonstrated to participate in the β-catenin/SOX signaling pathway, which regulated the chondrogenesis of hADSc. Taken together, these results and the established methodology contribute to a better understanding of cell morphology and roughness. Further, the data provide thermodynamic and kinetic insight into the integrin β1 ligand-binding process, at the single-molecule level. This AFM method will be useful for investigation of signaling pathways in living hADSc during chondrogenesis. Changes in the cellular nanostructure, as well as structure of the membrane, and the binding probability of transmembrane proteins are useful markers to evaluate chondrogenic differentiation mechanisms. This AFM method can be used to understand the mechanism of mesenchymal stem cell differentiation in tissue engineering and will be useful for an enhanced understanding of mesenchymal stem cell chondrogenic differentiation.