BMC(박테리아 미세구획)는 원핵생물 왕국에 널리 퍼져 있는 단백질성 자가 조립 소기관입니다. 다면체 껍질을 사용하여 주요 대사 효소 및 경로를 분할함으로써 BMC는 탄소 동화, 병인 및 미생물 생태학에서 필수적인 역할을 합니다. BMC 쉘은 정의된 아키텍처를 형성하기 위해 자가 조립되는 여러 단백질 동족체로 구성됩니다. 새로운 나노생물반응기 및 분자 스캐폴드를 개발하기 위해 BMC를 엔지니어링하는 데 엄청난 관심이 있습니다. 여기에서 우리는 고속 원자력 현미경(HS-AFM)을 사용하여 다양한 pH 및 염 조건에서 BMC 쉘 단백질의 형성 및 자가 조립 역학의 정량적 특성을 보고합니다. 우리는 400mM 염 농도가 쉘 헥사머에 의해 형성된 더 큰 단일층 쉘 패치를 생성하는 경향이 있고, 중성 pH에서 헥사머 자가 조립의 더 높은 동적 속도가 관찰되었음을 보여줍니다. 우리는 또한 6량체 어셈블리에서 섬유와 같은 어레이에 이르기까지 쉘 단백질의 가변성을 시각화합니다. 이 연구는 미세 환경 변화에 대한 응답으로 BMC 단백질 자체 조립의 안정성과 가변성에 대한 우리의 지식을 발전시키며, 이는 자가 조립 및 구조적 견고성을 리모델링할 수 있는 능력을 갖춘 합성 BMC 구조의 합리적인 설계 및 구성을 알려줄 것입니다. 또한 생명 공학 응용 분야에서 자가 조립 및 BMC 기반 나노 구조 형성을 정량적으로 평가하기 위한 강력한 도구 상자를 제공합니다.
소개
BMC(박테리아 미세구획)는 구조적으로 바이러스 캡시드와 유사한 단백질성 소기관으로 박테리아의 세포질을 분할합니다[1]. 그것들은 박테리아 문 사이에 널리 퍼져 있으며[2], 진핵생물에서 발견되는 막 결합 소기관이 없을 때 박테리아가 주요 대사 경로를 구획화할 수 있도록 합니다[3, 4]. BMC는 관강 효소 코어를 캡슐화하는 반투과성 단백질 껍질에 의해 형성됩니다. 쉘은 BMC-H(Pfam00936 도메인 포함), BMC-T(2개의 Pfam00936 도메인 포함) 및 BMC-P(Pfam03319 도메인 포함)를 포함하는 세 가지 유형의 구조적 단백질 구성요소로 구성됩니다[5,6,7, 8,9]. 쉘의 주요 구성 요소는 BMC-H로 볼록 및 오목 표면이 있는 6량체로 나타나며 오목면이 바깥쪽을 향하도록 쉘 패싯을 타일링합니다[10](그림 1). BMC-P는 정이십면체 모양의 꼭짓점을 덮도록 제안된 오량체를 형성하고 BMC-T는 껍질 투과성을 담당하는 것으로 추정되는 껍질 면에 위치한 유사 육량체를 형성합니다.
<그림>
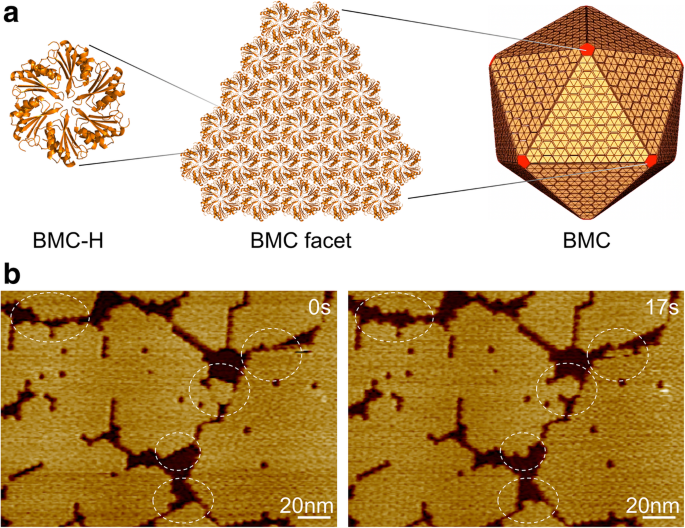
박테리아 미세 구획, 껍질 조직 및 자가 조립. 아 수백 개의 BMC 껍질 단백질 동족체가 자가 조립되어 20면체 단백질 소기관을 형성합니다. 노란색의 BMC-H 단백질은 면을 형성하고 빨간색의 BMC-P 단백질은 정점을 차지합니다. ㄴ Hoch_5815 BMC-H 헥사머로 구성된 쉘 패싯의 AFM 지형도. HS-AFM을 사용하여 몇 초 이내에 동적 이벤트(원)를 관찰했습니다.
특정 단백질-단백질 상호작용은 BMC 단백질의 자가 조립을 보장하여 대사 기능을 수행하기 위해 고도로 정의된 아키텍처를 형성합니다. 껍데기 단백질 사이의 측면 상호작용은 20면체 껍데기의 자가 조립 특성을 결정하는 주요 요인으로 간주됩니다[10]. BMC-H 동족체는 2차원 시트[11, 12], 나노튜브[13,14,15,16,17] 및 필라멘트 구조[15, 18,19,20]를 포함한 다양한 모양을 형성할 수 있음이 관찰되었습니다. .
자가 조립, 선택적 투과성 및 자연적으로 발생하는 세포 소기관의 효소 캡슐화 특성을 기반으로 BMC는 대사 효소를 둘러싸고 새로운 분자 지지체 생성을 통한 나노 규모 생물 반응기의 생체 영감 구성을 포함하여 생물 공학에서 큰 잠재력을 가진 이상적인 시스템으로 간주되었습니다. 새로운 기능 [21,22,23,24,25,26]. 그러나 BMC 구조가 얼마나 안정적인지, BMC 단백질 응집체의 자가 조립 및 형성을 효과적으로 조작 및 평가하는 방법과 같은 몇 가지 주요 문제는 BMC 생명공학에서 해결해야 할 문제로 남아 있습니다. BMC 쉘 및 전체 BMC의 구조 및 조립에 대한 조사는 X선 결정학, 전자 현미경(EM), 형광 현미경 및 동적 광산란(DSL)을 사용하여 수행되었습니다[10, 11, 16, 22, 27,28 ,29,30,31]. 최근에 우리는 고속 원자현미경(HS-AFM)을 활용하여 BMC-H 단백질의 동적 자가 조립 과정의 첫 번째 시각화를 수행했습니다[12].
이 작업에서 우리는 HS-AFM을 사용하여 다양한 pH 및 이온 조건에서 BMC-H 패치의 구조적 역학을 모니터링합니다. 이는 BMC 껍질 단백질 어셈블리의 변조에 대한 통찰력을 제공하고 분자 분해능에서 정량적 평가를 위한 강력한 도구를 제공합니다 , BMC 껍질 단백질 자가 조립의 안정성과 가변성에 대해.
방법
샘플 준비
Haliangium ocraceum에서 정제된 BMC-H 단백질(Hoch_5815) Kerfeld 박사(로렌스 버클리 국립 연구소)가 친절하게 제공했습니다. 완충액 교환의 경우 샘플을 ~ 80 mg mL
−1
로 보관하십시오. 트리스 완충액(50 mM Tris-HCl, pH 7.8, 100 mM KCl, 10 mM MgCl2 ) 0.5 mg mL
−1
로 희석했습니다. AFM 이미징 전에 원하는 버퍼를 사용합니다(추가 파일 1:그림 S1). 대조 완충액은 50mM Tris-HCl(pH 7.8) 및 10 mM MgCl2입니다. .
원자력 현미경
운모 및 AFM 이미징에서 샘플 흡수를 위해 원하는 버퍼를 사용했습니다. 운모에서 5분 흡수 후 Hoch_5815를 원하는 완충액으로 헹구어 고정된 단백질을 제거한 다음 AFM을 사용하여 이미지화했습니다(추가 파일 1:그림 S1). HS-AFM 이미지는 ULTRA Speed 2.8 μm 스캐너와 Ultra-Short Cantilever USC-0.3 MHz 프로브(NanoWorld)가 장착된 JPK NanoWizard ULTRA 속도 원자현미경을 사용하여 AC 모드의 솔루션에서 30 또는 40 Hz에서 캡처되었습니다. AFM 이미징 동안 ~ 100 피코뉴턴의 최소 하중을 적용하여 단백질 어셈블리의 교란을 줄였습니다[12, 32,33,34,35,36].
이미지 처리 및 분석
이미지 분석은 처음에 JPK SPM 데이터 처리(JPK)를 사용하여 수행되었습니다. HS-AFM 이미지 분석은 앞서 설명한 대로 Image SXM(http://www.ImageSXM.org.uk)에서 사용자 정의 매크로를 사용하여 수행되었습니다[12]. Hoch_5815 패치의 크기를 분석하기 위해 30 Hz 스캔 속도에서 캡처된 512 × 512 픽셀의 이미지를 평면화하여 XY 기울기와 Z 임계값을 제거한 다음 단백질이 아닌 단백질을 표시하기 위해 이진 변환을 수행했습니다. 입자 분석을 사용하여 이러한 이진 이미지에서 단백질의 표면적을 계산했습니다. 개별 패치와 인접 패치를 식별하기 위해 패치는> 3픽셀(~ 2 nm)로 구분된 개체로 정의되었습니다. 초기 테스트에서는 더 많은 수의 픽셀이 설정되면 인접한 패치가 단일 연속 패치로 계산될 수 있는 반면 더 작은 픽셀 수를 사용하면 패치의 개별 헥사머 사이의 간격이 패치 간의 경계로 잘못 계산될 수 있음을 보여주었습니다. 단백질 역학을 분석하기 위해 40Hz 스캐닝 속도로 캡처된 256 × 256 픽셀의 이미지 시리즈를 분석하여 프레임당 약 6.4 s의 시간 해상도를 제공했습니다. 차이 AFM 이미지를 표시하기 위해 시리즈의 이전 이미지에서 이진 이미지를 뺍니다. 차이 이미지의 입자 분석을 사용하여 조립 및 분해된 단백질의 면적을 계산했습니다. 동적 비율을 계산하는 데 사용되는 방정식은 다음과 같습니다.
여기서 N 는 임계값 차이 이미지의 흰색 픽셀과 검은색 픽셀의 합을 해당 축척의 단일 헥사머에 해당하는 픽셀 수로 나눈 값을 나타냅니다(추가 파일 1:그림 S3, 그림 S5). 데이터는 평균 ± 표준편차(SD)로 표시됩니다. 지정된 대로 다변수 ANOVA 또는 양방향 ANOVA를 사용하여 통계 분석을 수행했습니다.
<섹션 데이터-제목="결과">
결과
우리는 점액박테리움 Haliangium ocraceum의 BMC-H 단백질(Hoch_5815)을 사용했습니다. , Escherichia coli에서 표현된 6중 대칭을 갖는 6량체로 특징지어집니다[12]. Hoch_5815 헥사머는 20면체 BMC 아키텍처의 기본 구조 구성 요소를 나타내는 두 번째 시간 척도에서 단일 레이어 시트를 형성하기 위해 자가 조립될 수 있습니다(그림 1a). HS-AFM 이미징을 사용하면 시트 조각의 동적 조립 및 조직적 유연성을 시각화할 수 있고(그림 1b) 개발된 이미징 분석을 사용하여 패치 크기와 BMC-H 단백질의 동적 속도를 정량적으로 추정할 수 있습니다("방법" 섹션 참조).
pH 변화에 대한 반응
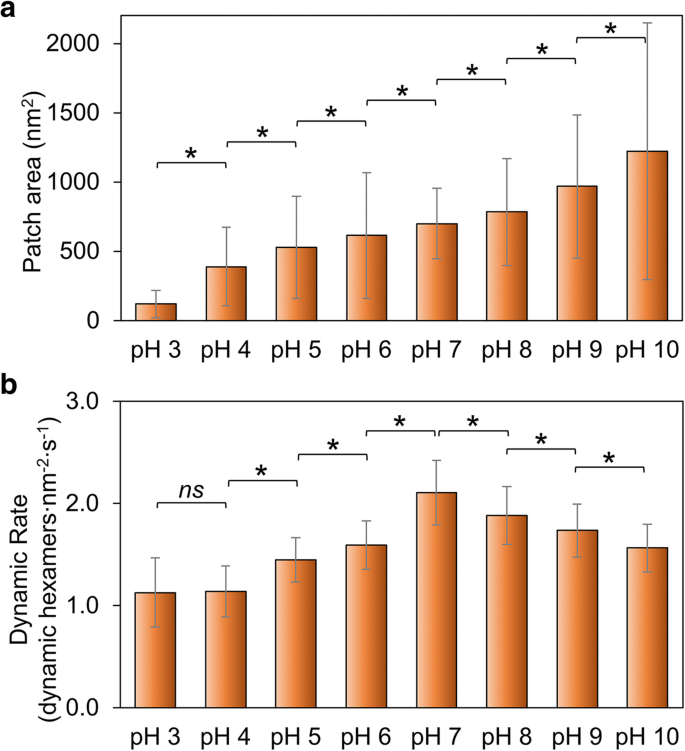
우리는 Hoch_5815의 자가 조립 능력의 전반적인 지표로 패치 크기의 변화를 측정했습니다. 패치 크기는 pH가 3에서 10으로 증가함에 따라 증가합니다(그림 2a, 추가 파일 1:그림 S2, 표 S1). 이는 높은 pH가 낮은 pH 조건보다 Hoch_5815 단백질의 자가 조립에 더 유리함을 시사합니다. 이것은 pH 6에서 불용성인 것으로 밝혀진 RmmH 단백질의 조립 거동과 다소 구별되며, pH 8에서는 정렬된 나노튜브 배열을 형성하고 pH 10에서는 분해되는 경향이 있습니다[13]. 또한 HOCH_5815 자체 조립의 높은 수준의 구조적 변동성을 관찰했습니다(그림 2a, 추가 파일 1:그림 S2에서 큰 SD로 표시됨).
<그림>
Hoch_5815의 자가 조립에 대한 환경 pH의 영향. 아 AFM에 의해 결정된 Hoch_5815의 개별 패치의 평균 표면적(n =50) (추가 파일 1:그림 S2). ㄴ HS-AFM(n =50). *p <0.05, ns 중요하지 않음(다변량 ANOVA)
쉘 시트에서 Hoch_5815 단백질의 자가 조립에 대한 AFM 영상은 쉘 시트의 형성이 6량체의 조립 및 분해의 조합에 기인한다는 것을 밝혀냈습니다[12]. Hoch_5815 단백질-단백질 상호작용의 안정성을 조사하기 위해 다양한 pH(추가 파일 1:표 S2)에서 Hoch_5815 자가 조립 역학 및 동적 이벤트의 속도를 추가로 조사했습니다. 자가 조립 역학의 비율은 pH 7에서 가장 높고 산성 및 알칼리성 조건 모두에서 감소합니다(그림 2b, 추가 파일 1:그림 S3). 특히 pH 7에서 pH 6으로 산성 조건에서 급격히 감소하고 그림 2b와 같이 pH 4와 pH 3 사이에서 상대적으로 일정하게 나타납니다.
pH는 hexamer-hexamer 계면에 위치한 아미노산 잔기의 정전기적 특성에 큰 영향을 미칠 가능성이 있습니다. 산성 조건에서 관찰된 감소된 역학과 더 작은 크기의 쉘 패치는 Hoch_5815가 감소된 자가 조립 능력을 갖고 있음을 보여줍니다. 알칼리성 조건에서 관찰된 감소된 역학 및 더 큰 크기의 쉘 패치는 안정적인 6량체-헥사머 상호 작용을 시사하는 반면 Hoch_5815 6량체의 증가된 역학은 중성 pH 조건에서 유연한 6량체-헥사머 상호 작용을 의미합니다.
염류 농도 변화에 대한 대응
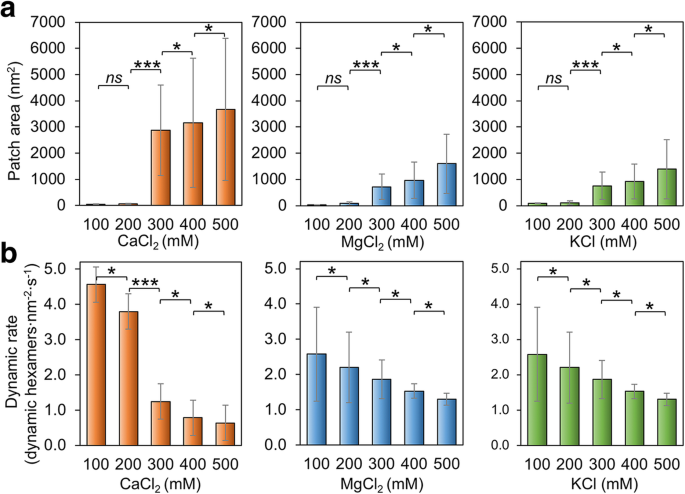
또한 버퍼의 염 농도가 Hoch_5815 조립에 영향을 미치는지 확인했습니다. MgCl2의 낮은 농도(100–200 mM)에서 , CaCl2 및 KCl, Hoch_5815 단백질은 더 높은 농도(300–500 mM)에서 조립된 것보다 상대적으로 작은 패치를 형성합니다(그림 3a, 추가 파일 1:그림 S4). 500 mM에서 이중 또는 다중 레이어 Hoch_5815 시트를 관찰했습니다(추가 파일 1:그림 S4). 이러한 관찰은 더 높은 이온 강도가 탄소 동화를 위한 카르복시솜의 껍질 단백질인 CcmK에 의해 보다 광범위하고 잘 정렬된 2D 결정의 형성을 촉진할 수 있다는 이전의 발견과 일치합니다[37]. 그러나 RmmH에 의해 형성된 고도로 정렬된 나노튜브는 NaCl 농도가 50에서 500 mM으로 증가할 때 분해되었으며[13], 이는 쉘 6량체에 의한 평평한 시트 및 관 모양의 형성을 매개하는 잠재적으로 다른 메커니즘을 나타냅니다.
<그림>
Hoch_5815의 자가 조립에 대한 염 농도의 영향. 아 100–500 mM CaCl2 범위에서 AFM으로 측정한 평균 패치 면적 , MgCl2 , 및 KCl(n =50). 염 농도의 증가는 패치 크기를 증가시켰습니다. 패치 면적의 상당한 변화가 200과 300 mM 사이에서 관찰되었습니다(***p <0.001, *p <0.05, ns 유의하지 않음, 양방향 ANOVA). ㄴ 100–500 mM CaCl2 범위에서 고속 AFM 이미지 시리즈에서 결정된 동적 이벤트의 평균 비율 , MgCl2 , 및 KCl(n =50). 염 농도의 100 mM 변화는 동적 이벤트의 비율에 상당한 변화를 가져왔습니다(***p <0.001, *p <0.05, ns 유의하지 않음, 양방향 ANOVA)
또한 MgCl2의 변화로 인한 Hoch_5815 자가조립의 변형 및 KCl 농도는 비교적 유사합니다. 대조적으로, 패치 크기의 변화는 CaCl2일 때 가장 두드러집니다(최대 3000배 증가). 농도가 200에서 300 mM으로 증가했으며(그림 3a), Hoch_5815 자가 조립의 CaCl2에 대한 더 높은 감도를 나타냅니다. MgCl2보다 또는 KCl.
Hoch_5815 자체 조립의 동적 속도는 완충염 농도의 변화에도 영향을 받습니다. MgCl2의 증가 , CaCl2 , 또는 KCl 농도는 Hoch_5815 동적 속도의 감소를 초래할 수 있습니다(그림 3b, 추가 파일 1:그림 S5). 더 높은 염 농도에서 관찰된 패치 크기의 증가를 감안할 때(그림 3a), Hoch_5815 헥사머 사이의 측면 상호작용이 높은 염 농도에서 더 안정한 것으로 보입니다. CaCl2의 변화 농도는 더 확연한 반응을 보였고 200과 300 mM 사이의 동적 이벤트 비율에 상당한 변화가 있었습니다(그림 3b). 반면 MgCl2의 변화에 대한 반응 및 KCl은 패치 크기의 변화와 일치하여 비교적 유사합니다(그림 3a). 흥미롭게도, 조립 이벤트 대 분해 이벤트의 가장 높은 비율은 MgCl2의 400 mM에서 관찰되었습니다. , CaCl2 또는 KCl(추가 파일 1:표 S2). 이로 인해 400 mM 염에서 크고 안정적인 단일층 Hoch_5815 어셈블리가 형성되었습니다(추가 파일 1:그림 S4). 500 mM에서 관찰된 이중층 어셈블리도 안정적이며 헥사머 이동 속도가 낮습니다.
BMC-H 단백질 조립의 유연성
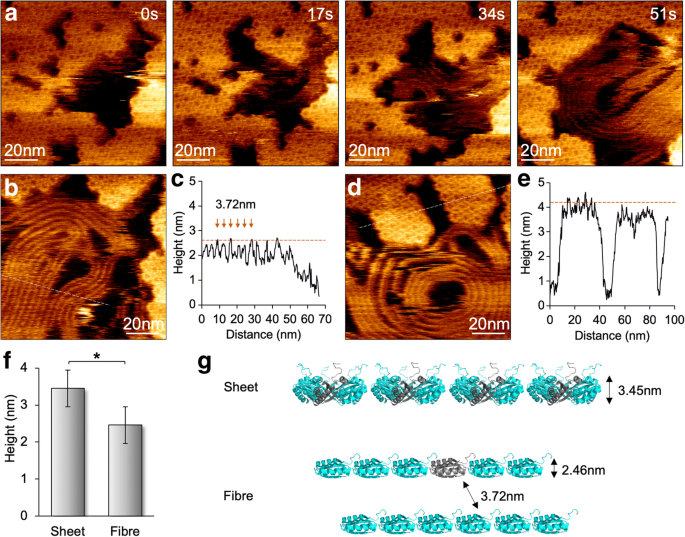
스캐닝 힘을 100pN으로 줄임으로써 BMC 단백질 어셈블리에 대한 AFM 팁 스캐닝의 영향을 최소화하고 개별 헥사머의 분자 분해능 AFM 이미지를 얻었습니다(그림 4). 조립 및 분해 이벤트 모두 동일한 보기에서 볼 수 있으며 팁 스캐닝 아티팩트 대신 BMC 쉘 조립의 동적 특성을 확인할 수 있습니다[12]. HS-AFM 영상은 또한 Hoch_5815 단백질 응집체의 가변성을 보여주었습니다. 10 mM MgCl2만 존재하는 pH 7.5에서 샘플을 이미징할 때 놀랍게도, 우리는 두 번째 시간 척도에서 Hoch_5815 헥사머의 분해와 함께 섬유 유사 구조의 형성을 때때로 관찰했습니다(그림 4a). 이러한 섬유와 같은 구조는 쉘 헥사머에 의해 조립된 나노튜브 번들과 유사하게 병렬로 조밀하게 패킹될 수 있습니다[13,14,15,16]. 그러나 두 섬유 사이의 공간은 3.72 ± 0.31 nm(n =30)이고 평균 높이는 2.46 ± 0.22 nm(n)입니다. =30), Hoch_5815 6량체(3.45 ± 0.16 nm, n)에 의해 형성된 쉘 시트보다 작음 =25) (그림 4b–f). 이러한 섬유 구조는 이미징 중에 상당히 유연하고 동적이며 다양한 크기의 직선 또는 코일 구조를 표시할 수 있습니다. Hoch_5815 헥사머의 분해가 수반되는 섬유 구조의 외관과 단층 헥사머 시트와 비교하여 감소된 높이를 감안할 때 이러한 섬유 유사 구조는 헥사머에서 분해된 개별 Hoch_5815 펩티드에 의해 형성되는 것으로 추측됩니다(그림 4g). . 특정 완충 조건(예:낮은 이온 강도)에서 기질 흡수는 기질 표면에 Hoch_5815 펩타이드의 알파 나선 면이 부착되고 인접한 펩타이드와 펩타이드의 선형 결합으로 이어질 가능성이 있습니다. 6량체 내 상호 작용이 강할 가능성이 높다[5]. 껍질 단백질 응집의 변동성을 뒷받침하는 자세한 메커니즘은 더 자세히 설명해야 합니다.
<그림>
HS-AFM에서 쉘 시트 어셈블리와 함께 섬유 구조의 형성 및 역학. 아 AFM 이미지 시리즈에서 볼 수 있듯이 Hoch_5815 헥사머로 구성된 쉘 시트의 분해 중 섬유 유사 구조의 모양. ㄴ 섬유 구조의 AFM 지형도. ㄷ 횡단면 분석(패널 b의 파선 ) 3.72 ± 0.31 nm(n =30) 인접한 섬유 구조 사이, 평균 높이는 2.46 ± 0.22 nm(n =30). d Hoch_5815 헥사머로 구성된 쉘 패치의 AFM 지형도. 이 횡단면 분석(패널 d의 파선 ) Hoch_5815 헥사머의 평균 높이는 3.45 ± 0.16 nm(n =25). 에 섬유와 같은 구조는 Hoch_5815 헥사머(*p <0.05, 양방향 ANOVA). 지 일련의 Hoch_5815 단량체를 나타내는 섬유와 같은 구조의 제안된 구성 및 형성
토론
BMC는 자가 조립하여 고차 구조를 형성하는 수백 개의 단백질로 구성됩니다. 수많은 단백질 동족체로 구성된 BMC 껍질은 단백질 자기 조립 및 상호 작용을 연구하는 데 이상적인 시스템입니다. AFM은 생물학적 시스템의 생리적 역할과 매우 관련이 있는 생체막 조직, 단백질 조립 및 물리적 상호작용을 분석하기 위한 강력한 기술로서[32, 35, 38, 39], 원자현미경은 조직 및 자가 조립 역학을 시각화하는 데 활용되었습니다. BMC 껍질 단백질과 BMC 구조의 구조 및 기계적 특징 [12, 30, 31, 40,41,42]. 이 작업은 우리가 아는 한 AFM을 사용하여 환경 변화에 반응하여 2차원 시트 형성에서 BMC 껍질 단백질의 자가 조립 역학의 첫 번째 정량적 결정을 나타냅니다. 결과는 BMC-H 단백질 자가 조립의 고유한 가변성과 환경 의존성을 강조합니다. EM 및 DSL과 비교하여 AFM은 분자 세부 사항을 통해 실시간으로 BMC 단백질 자가 조립의 동적 작용을 모니터링하는 데 큰 잠재력을 보여줍니다.
단백질-단백질 상호작용은 BMC 껍질을 형성하고 형성하는 데 매우 중요합니다[10]. 단백질 농도는 또한 껍데기 형성을 주도하는 중요한 요소로 문서화되어 있습니다[41, 43]. 또한, 시험관 내 용해도 연구는 용액의 pH와 이온 강도가 BMC의 구조적 안정성[17, 27]과 2차원 시트 형성에서 BMC 껍질 단백질의 조립 거동에 영향을 줄 수 있음을 보여주었습니다[37, 41 ], 나노튜브[13, 17] 및 나노케이지[28]는 바이러스 캡시드 어셈블리에 미치는 영향을 연상시킵니다[44, 45]. 우리는 또한 pH> 10 및 <3 또는 염 농도 <10 mM 또는> 600 mM일 때 단백질 침전 및 패치가 형성되지 않음을 발견했습니다(미공개 데이터). 여기서 우리는 어셈블리 경향과 역학이 pH와 염 농도에 의존한다는 것을 추가로 보여주었습니다. 껍질 단백질은 광범위한 pH에서 자가 조립할 수 있지만 중성 pH 환경은 조립 역학을 향상시킬 수 있는 것으로 보입니다(그림 2b). 농도가 300 mM 이상인 양이온은 2차원 시트의 형성을 촉진하는 것으로 밝혀졌습니다. 400 mM 양이온은 크고 안정적인 단층 시트의 형성에 바람직한 것으로 보입니다(그림 3). 이러한 조건은 세균 세포의 세포질 조건과 일치하며 생리학적으로 관련이 있습니다. 예를 들어, 생리학적으로 가장 적절한 조건에서 E의 pH. 대장균 세포질은 약 7.4~7.8[46]이고 이온 농도는 약 100~400 mM이며, 이는 단백질 상호 작용, 단백질-리간드 결합, 신호 전달, 막 정전기 전위 유지 및 막을 가로지르는 단백질 구배에 필수적입니다[47, 48]. 샘플과 운모 기질 사이의 상호 작용이 BMC 단백질의 자가 조립에 어떻게 영향을 미치는지 더 조사해야 하지만 AFM 이미징은 환경 변화에 대한 응답으로 BMC 단백질 자가 조립의 동적 변화를 정량적으로 분석할 수 있는 기회를 제공합니다.
여기에 설명된 껍질 조각의 형성에서 BMC 단백질의 환경 의존적 어셈블리 역학은 전체 BMC 형성에서의 행동을 나타낼 수 있습니다. 사실, 3D BMC 구조는 자연에서 설계된 동적으로 유지되는 소기관인 것처럼 보입니다. BMC는 주목할만한 구조적 유연성과 이질성을 나타냅니다. AFM 나노인덴테이션[30]에 의해 결정된 BMC 쉘 구조의 기계적 부드러움과 전산 시뮬레이션[49]에 의해 밝혀진 BMC 어셈블리의 비평형 역학은 BMC와 강력한 바이러스 어셈블리 간의 차이점을 강조했습니다. 마찬가지로, 카르복시솜의 생합성은 빛 및 샤페론과 상관관계가 있는 것으로 밝혀졌습니다[50, 51]. 아주 최근에, CcmK3 및 CcmK4가 헤테로헥사머를 형성하고 pH 의존적 방식으로 카르복시 껍질을 덮을 수 있으며, 카르복시 껍질 투과성과 CO2를 조절하는 수단을 제공할 수 있음이 밝혀졌습니다. 매우 역동적인 미세 환경에서의 동화 [52]. 용액의 환경 조건이 BMC 단백질의 열역학적 조립에 어떻게 영향을 미치는지에 대한 정확한 메커니즘은 예를 들어 실험 연구와 컴퓨터 시뮬레이션의 조합을 사용하여 조사해야 합니다.
BMC 구조의 자가 조립을 감안할 때 BMC 엔지니어링 및 새로운 BMC 기반 나노바이오리액터, 분자 스캐폴드 및 생명 공학 응용 분야의 생체 재료 설계(예:세포 대사, 효소 캡슐화, 분자 전달 및 치료 향상)에 상당한 관심이 있습니다. . 환경 변화에 대응하는 BMC의 구조적 탄력성과 가변성에 대한 고급 지식은 이종 호스트, 즉 E. 대장균 또는 식물 [31, 53, 54]뿐만 아니라 2D 나노 물질의 형성과 BMC 껍질 기반 단백질 케이지의 개폐를 조절하여 기능 조절 및 표적 분자 전달을 촉진하는 길을 열어줍니다. 이전에 우리는 쉘 단백질의 경계면에서 특정 접촉을 조작하기 위해 유전자 변형 접근 방식을 사용하는 가능성과 자체 조립 동작을 입증했습니다[12]. 이 연구는 다양한 환경에서 BMC 쉘 자체 조립을 평가하고 조작하기 위한 도구 상자를 강화합니다.
결론
요약하면, 우리는 HS-AFM을 이용하여 다양한 pH 및 염 조건에서 BMC 쉘 단백질 자가 조립의 정량적 조사를 수행했습니다. 쉘 헥사머의 더 큰 단일층 패치의 형성은 400-mM 염 농도에서 촉진되는 것으로 나타났으며 중성 pH는 헥사머 자가 조립의 더 높은 동적 속도를 초래했습니다. 6량체 어셈블리에서 섬유와 같은 어레이로 쉘 단백질의 조직적 전환도 시각화되었습니다. 이 연구는 환경 조건이 BMC 껍질 단백질의 조직과 자가 조립을 결정하는 데 중요한 역할을 한다는 것을 보여주었습니다.