다중벽 탄소 나노튜브 유발 토마토 측근 형성의 기초가 되는 새로운 메커니즘:산화질소의 관여
초록
많은 연구에서 다중벽 탄소나노튜브(MWCNT)가 식물에 유독하다는 것이 밝혀졌습니다. 그러나 MWCNT가 다양한 환경 신호에 대한 뿌리 시스템의 적응성의 중요한 구성 요소인 측면 뿌리(LR) 형성에 영향을 미치는지 여부는 여전히 논란의 여지가 있습니다. 이 보고서에서 우리는 MWCNT가 토마토 묘목 뿌리에 들어갈 수 있음을 발견했습니다. MWCNT를 사용한 투여는 대략 용량 의존적 방식으로 토마토 LR 형성을 촉진했습니다. 내인성 산화질소(NO) 생성은 MWCNTs에 의해 유발되었으며, 이는 Greiss 시약 방법, 전자 상자성 공명(EPR) 및 레이저 주사 공초점 현미경(LSCM)에 의해 확인되었으며 NO의 제거제와 함께 확인되었습니다. MWCNT에 의해 유발된 NO 합성과 LR 형성은 스캐빈저로 내인성 NO를 제거함으로써 분명히 차단되었기 때문에 LR 발달의 유도에서 MWCNT와 NO 사이에는 인과 관계가 존재합니다. NO 생성 효소인 NR(nitrate reductase)의 활성은 MWCNT에 반응하여 증가했습니다. NR의 텅스텐산 억제는 NO 생성을 손상시켰을 뿐만 아니라 MWCNT에 의해 유발된 LR 형성을 없앴습니다. N 추가
지
-nitro-l-arginine methyl ester(l-NAME), 포유류의 NOS(nitric oxide synthase) 유사 효소 억제제는 LR 형성에 영향을 미치지 못했습니다. 종합적으로, 우리는 NO가 적어도 부분적으로 NR을 통해 LR 발달의 MWCNT 제어에서 다운스트림 신호 분자로 작용할 수 있다고 제안했습니다.
소개
탄소나노튜브의 생물학적 및 생물의학적 응용은 많다[1, 2]. 세포막을 쉽게 투과할 수 있는 독특한 능력으로 인해 탄소나노튜브의 생물학적 안전성은 항상 논쟁의 주제입니다[3, 4]. 한편, 탄소나노튜브의 생산 및 사용이 빠르게 증가하고 있기 때문에 인간과 포유류, 최근에는 식물에서 세포독성의 상세한 기전을 규명하는 것이 중요해지고 있다[3,4,5,6,7,8,9]. 식물과 그 군집은 인간과 환경에 매우 중요하며 생물고체 비료 시비를 통해 토양에 축적되기 때문에 탄소나노튜브 노출의 위험이 있다는 것은 잘 알려져 있습니다[6, 10, 11]. 탄소나노튜브의 중요한 구성요소로서, 그래핀의 다중 압연층으로 구성된 다중벽 탄소나노튜브(MWCNT)의 독성이 널리 연구되어 왔다. 포유류에 대한 연구에 따르면 MWCNT와 단일벽 탄소 나노튜브 모두에 노출되면 인간 각질세포와 A549 세포에서 산화적 손상과 NF-κB 활성화가 유도되는 것으로 나타났습니다[9, 12]. MWCNT와 단일벽 탄소나노튜브는 원형질막과 융합하여 지질 과산화와 산화 스트레스를 통해 세포 손상을 일으킬 수 있다[9, 11, 13, 14]. MWCNT에 의해 유발된 세포 독성 및 산화 스트레스, 그리고 적당한 염증 반응이 인간 제대 정맥 내피 세포에서 관찰되었습니다[15]. 이전 연구에서는 붉은 시금치에 있는 MWCNT의 1차 독성이 주로 활성 산소종(ROS) 과잉 생산에서 유래했으며, 아스코르브산을 보충하면 독성 효과를 역전시킬 수 있다고 제안했습니다[7]. 이러한 의미에서 MWCNT는 동물이나 식물에서 유기체에 새로운 스트레스 요인으로 간주됩니다.
뿌리 구조의 중요한 결정 요인인 측면 뿌리(LR) 형성은 다양한 스트레스에 대한 적응적 반응의 지표로 간주되어 왔습니다[16]. 고등 식물에서 LR의 형성은 식물 호르몬과 수분 가용성, 영양소, 저산소증 및 중금속 스트레스와 같은 비생물적 스트레스를 포함한 광범위한 환경 신호의 영향을 받습니다[17,18,19]. 한편, LR의 형성은 물리적 지지체로서 작용할 뿐만 아니라 식물 성장 및 발달을 위한 물 및 영양소 흡수에 기여한다는 충분한 증거가 확인되었습니다[19,20,21]. 다양한 환경적 단서는 LR 형성 촉진 및 뿌리 신장 억제를 포함하여 몇 가지 특정 스트레스 유발 형태 반응(SIMR) 표현형을 유발할 수 있습니다. [17] LR 형성의 조절은 또한 옥신, 그리고 옥신에 대한 반응으로 세포주기 조절 유전자의 활성화가 제안되었다[19, 22]. 한편, 오이, 토마토, 대두 및 유채 식물에서 옥신 유발 뿌리 기관 형성에 일부 소분자의 관여가 확인되었습니다[23,24,25,26,27]. 이러한 작은 분자에는 과산화수소(H2 O2 ), 산화질소(NO), 일산화탄소(CO) 및 수소 가스(H2 ).
이 중 자유 라디칼 가스인 NO는 식물에서 여러 생리학적 기능을 갖는 것으로 밝혀졌습니다[28, 29]. 스트레스에 대한 식물 적응의 향상 외에도 NO의 기능에는 뿌리 모발 발달 촉진, 우발적 발근 및 측근 형성이 포함되지만[30,31,32,33], 위에서 언급한 프로세스는 여전히 파악하기 어렵습니다. 동물에서 l-아르기닌으로부터의 NO 합성은 헴 함유 효소인 산화질소 합성효소(NOS)에 의해 촉매됩니다[34]. NOS 효소를 암호화하는 유전자는 식물에서 확인되지 않았지만 포유동물의 NOS 유사 활성이 널리 검출되고[35, 36], N과 같은 포유동물 NOS의 억제제가
지
-nitro-l-arginine methyl ester hydrochloride(l-NAME)는 식물에서 NO 생성을 억제할 수 있습니다[25, 33, 36,37,38,39]. 중요하게도, 풍부한 유전적 증거는 NO가 식물에서 질소 대사를 담당하는 잘 알려진 효소인 질산염 환원효소(NR)에 의해 생성될 수 있음을 보여주었습니다[28]. NR 매개 NO 생산이 기공 폐쇄 및 저온 순응에 관여하는 것은 유전적으로 입증되었습니다[37, 38]. 우리의 이전 연구는 NR 의존적 NO 합성이 옥신 유도 수소 가스 매개 측근 형성에 관여한다는 것을 보여주었습니다[39].
지금까지 MWCNT[40,41,42,43], 금 나노입자(Au NP, [44]), 산화아연을 포함한 나노물질이 보충되었을 때 다양한 식물 종에서 LR 형성, 촉진 또는 억제에 대한 서로 다른 반응이 보고되었습니다. 나노입자(ZnO NP [45, 46];), 이산화티타늄 나노입자(TiO2 NP [46];) 및 산화 그래핀 (GO [47,48,49];) (표 1)이 있으며, 위의 반응에서 NO의 역할에 대한 확실한 증거를 제공한 연구는 아직 없습니다. 이 연구에서 Greiss 시약법, 레이저 주사 공초점 현미경(LSCM) 및 전자 상자성 공명(EPR) 분석에 의한 내인성 NO의 검출은 MWCNT 처리된 토마토 묘목에서 NO 수준이 증가된 것으로 나타났습니다. 그 후, LR 형성이 관찰되었다. 우리는 NR 및 포유류와 같은 NOS 활성을 억제하는 길항제와 NO 스캐빈저를 사용하여 내인성 NO 수준을 조작함으로써 MWCNT에 의해 촉발된 LR 형성에 NO의 관여를 추가로 연구합니다. 추가 실험은 NR 의존성 NO가 MWCNT에 대한 반응으로 LR 형성에 적어도 부분적으로 필수적일 수 있음을 보여주었습니다. 따라서 이 연구는 식물에서 나노물질의 생물학적 효과를 이해하기 위한 새로운 창을 엽니다.
자료 및 방법
화학물질
달리 명시되지 않는 한, 다른 모든 화학물질은 Sigma-Aldrich(St Louis, MO, USA)에서 구입했습니다. Sigma-Aldrich에서 구입한 MWCNT는 이전에 설명된 대로 특성화되었습니다[50]. MWCNT의 외경, 내경 및 길이는 각각 6–12 nm, 2.5–5 nm 및 1–9 μm였습니다. 초음파 처리 후, 얻어진 균질 콜로이드 현탁액을 멸균하여 사용하였다.
단일벽 탄소 나노튜브(SWCNT, XFS22, 순도> 95%, 직경 1–2 nm, 길이 5–30 μm, 특수 표면적> 1075 m
2
/g), 그래핀(XF001W, 순도 ~ 99%, 직경 0.5–5 μm, 두께 ~ 0.8 nm, 단층 비율 ~ 80%, BET 표면적 500~1000 m
2
/G; 전기 저항 ≤ 0.30 Ω.cm) 및 활성탄(AC, XFP06, 순도> 95%, 입자 크기 5 ± 1 μm, 기공 부피 1–1.2 cm
3
/g, 조리개 2.0–2.2 nm, 특수 표면적 ~ 1500–1700 m
2
/g).
또한, 나트륨 니트로프루시드(SNP)는 NO 방출 화합물로 사용되었습니다[30,31,32,33]. 2-(4-카르복시페닐)-4,4,5,5-테트라메틸이미다졸린-1-옥실-3-옥사이드 칼륨염(cPTIO)은 NO의 제거제로 간주되었다[51,52,53,54]. 텅스텐산염(Tg, NR의 억제제 [28, 33, 37, 55,56,57]) 및 N
지
-nitro-L-arginine methyl ester hydrochloride(NAME; 포유류 NOS 유사 효소의 억제제[25, 33, 36,37,38,39])도 적용되었습니다. 본 연구에서는 파일럿 실험에서 상기 화학물질의 농도를 결정하였고, 그 결과 유의미한 반응이 관찰되었다.
식물 재료 및 성장 조건 및 LR 형성의 결정
토마토(Solanum lycopersicum L.) 종자 "Jiangshu 14"는 중국 장쑤성 난징의 Jiangsu Agricultural Institutes에서 친절하게 공급되었습니다. 동일한 크기의 종자를 선별하여 25 ± 1 °C의 암실에서 증류수에 3 일간 발아시켰다. 그런 다음 2-3 mm의 근간을 가진 선택된 동일한 묘목을 표시된 농도의 MWCNT, 200 nM 1-나프틸아세트산(NAA; 잘 알려진 옥신), 0.1 mM SNP, 0.2 mM cPTIO, μM 텅스텐산염(Tg), 0.2 mM NAME 및 5 mg/mL 단일벽 탄소 나노튜브(SWCNT), 그래핀 및 활성탄(AC)을 포함한 기타 탄소 나노입자, 단독 또는 표시된 시점의 조합. 묘목을 조명 인큐베이터(25 ± 1 °C)에서 200 μmol m
−2
의 빛 강도로 재배했습니다. s
−1
14/10 h(밝음/어두움) 광주기에서.
처리 후 사진을 촬영하고 Image J 소프트웨어(http://rsb.info.nih.gov/ij/)를 사용하여 묘목당 출현한 측근의 수와 길이(> 1 mm)를 결정했습니다. , 58]. 이전에 설명한 바와 같이 측근 유도성 분절만 후속 분석에 사용되었습니다.
투과 전자 현미경에 의한 MWCNT 분포의 이미징
토마토 묘목 뿌리에서 MWCNT의 분포는 투과 전자 현미경(TEM; JEOL, JEM-200CX, Tokyo, Japan)을 사용하여 특성화되었습니다. TEM 분석을 위한 샘플 준비는 이전 프로토콜에 따랐습니다[59].
레이저 스캐닝 공초점 현미경에 의한 내인성 NO의 이미징
상당히 특정한 NO 형광 프로브 4-아미노-5-메틸아미노-2',7'-디플루오로플루오레세인 디아세테이트(DAF-FM DA)를 사용하여 NO 이미징을 수행했습니다. 프로브를 철저히 세척한 후 Zeiss LSM 710 공초점 현미경(Carl Zeiss, Oberkochen, Germany, 여기 488nm, NO 분석의 경우 500–530nm 방출)을 사용하여 이미지를 얻었습니다. 우리의 실험에서 20개의 개별 샘플을 무작위로 선택하여 처리당 측정했습니다. 사진은 동일한 결과를 나타냅니다.
Griess 시약 분석에 의해 결정된 콘텐츠 없음
이전에 설명한 방법[50]에 따라 Griess 시약 분석으로 NO 함량을 측정했습니다. 중요하게도, 식물의 농축된 질산염 및 아질산염 함량으로 인한 간섭을 피하기 위해 200μM cPTIO(NO 제거제)에서 30분 동안 사전 배양된 동일한 샘플을 빈 샘플로 간주했습니다. 30분 동안 Griess 시약을 첨가한 후, 540nm에서 흡광도를 기록하고, NaNO2의 표준 곡선과 비교하여 NO 함량을 결정했습니다. .
전자 상자성 공명(EPR)으로 NO 결정
우리의 이전 방법[39, 55, 60]에 따라 전자 상자성 공명(EPR)을 사용하여 NO 수준의 결정이 수행되었습니다. 유기 용매 층을 사용하여 다음 조건에서 Bruker A300 분광계(Bruker Instrument, Karlsruhe, Germany)에서 NO를 측정했습니다:실온; 마이크로파 주파수, 9.85 GHz; 마이크로파 전력, 63.49 mW; 및 변조 주파수, 100.00 kHz.
질산염 환원효소(NR) 활성 측정
NR 활성은 이전 방법에 따라 540 nm에서 분광광도계로 검출되었습니다[57]. 생성된 아질산염은 0.02%(1-나프틸)-에틸렌디아민 1 mL와 함께 3 M HCl 중 1%(w/v) 설페닐아미드 1 mL를 첨가하여 540 nm에서 분광광도계로 측정했습니다.
통계 분석
표시된 경우, 결과는 각각에 대해 3개의 생물학적 복제물이 있는 3개의 독립적인 실험의 평균값 ± SE로 표시되었습니다. 통계 분석은 SPSS Statistics 17.0 소프트웨어를 사용하여 수행되었습니다. 통계 분석을 위해 Duncan의 다중 검정(p <0.05)가 적절하게 선택되었습니다.
<섹션 데이터-제목="결과">
결과
MWCNT는 루트 세포에 들어갈 뿐만 아니라 LR 형성을 촉진합니다.
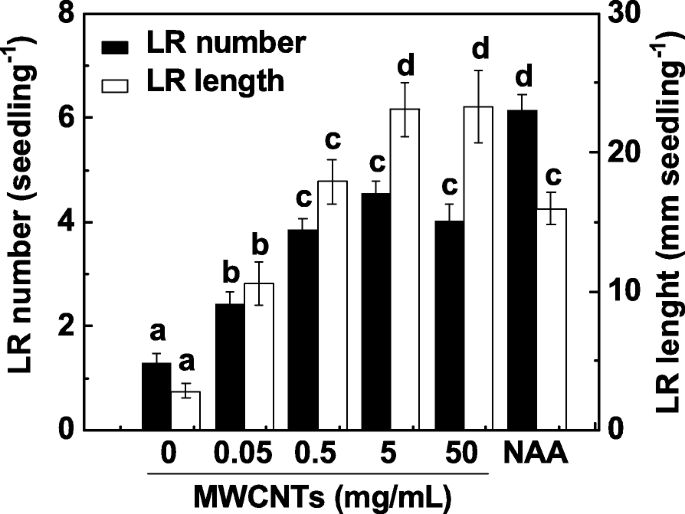
LR 형성은 루트 시스템 아키텍처의 주요 결정 요인입니다. LR 형성에 대한 MWCNT의 영향을 조사하기 위해 3일 된 토마토 묘목을 다양한 농도의 MWCNT(0.05, 0.5, 5 및 50 mg/mL)와 함께 3일 동안 배양했습니다. 1-나프틸아세트산(NAA)의 적용은 양성 대조군으로 간주되었다. 우리의 실험에서 LR 수와 길이는 모두 LR 형성의 두 가지 매개변수로 결정되었습니다. 도 1에 나타난 바와 같이, 대조군과 비교하여 외인성 MWCNT는 5 및 50 mg/mL에서 최대 효과와 함께 대략 용량 의존적 방식으로 LR 형성을 유의하게 유도하였다. 200 nM NAA를 투여했을 때도 유사한 유도성 반응이 관찰되었습니다. MWCNT의 비용과 LR 형성의 유도성 반응을 고려하여 다음 실험에 5 mg/mL MWCNT를 적용했습니다.
<그림>
MWCNT에 의해 유도된 토마토 LR 형성은 대략적으로 용량 의존적이었습니다. 3일 된 토마토 묘목은 각각 200 nM NAA와 표시된 농도의 MWCNT로 처리되었습니다. 그런 다음 묘목당 출현한 측근의 수와 길이(> 1 mm)를 처리 3일 후에 결정했습니다. 3개의 생물학적 복제물에 30(10 × 3)개의 식물이 있었고, 실험은 3번 수행되었다. 데이터는 평균 ± SE입니다. 각 실험 세트 내에서 동일한 문자로 표시된 막대는 p에서 크게 다르지 않았습니다. <0.05 Duncan의 다중 테스트에 따른 수준
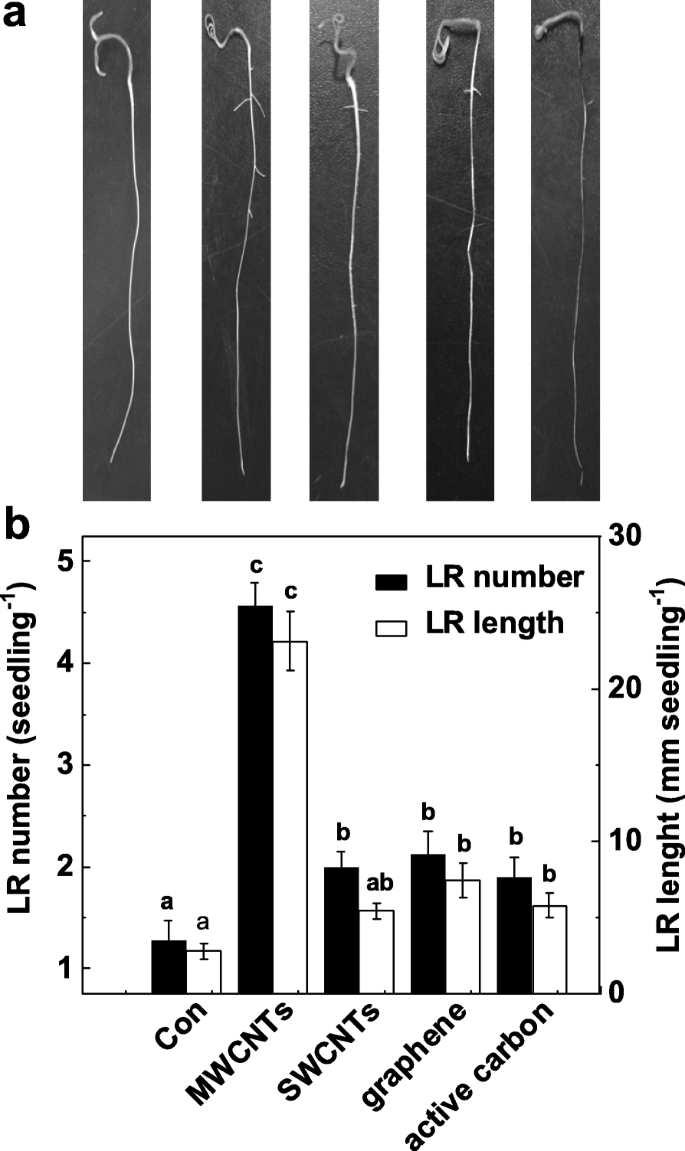
LR 형성 유도에서 MWCNT의 특정 기능을 검증하기 위해 우리는 MWCNT의 다른 동소체에도 이러한 유도 효과가 있는지 여부를 추가로 조사합니다. 도 2a에 나타난 바와 같이, 이러한 모든 탄소 나노물질은 싹 성장에 대한 독성 효과를 나타내었다(데이터는 나타내지 않음). 흥미롭게도 MWCNT, 단일벽 탄소 나노튜브(SWCNT), 그래핀 및 활성탄을 동일한 농도(5 mg/mL)로 적용하면 무화학 대조군 식물에 비해 LR 수와 길이가 다르게 증가할 수 있습니다. (그림 2b). 이들 화학물질 중 MWCNT 배양토마토 묘목에서 최대 유도성 반응이 발견되었다.
<그림>
다양한 탄소 나노 입자에 대한 측근 형성의 변화. 3일 된 토마토 묘목을 증류수(Con), 5 mg/mL MWCNT, 단일벽 탄소나노튜브(SWCNT), 그래핀 및 활성탄(AC)으로 각각 추가 3 일 동안 처리했습니다. 아 그런 다음 대표 사진을 찍었습니다. ㄴ 그런 다음 묘목당 출현한 측근의 수와 길이(> 1 mm)도 결정되었습니다. 스케일 바 =50 mm. 3개의 생물학적 복제물에 30(10 × 3)개의 식물이 있었고, 실험은 3번 수행되었다. 데이터는 평균 ± SE입니다. 각 실험 세트 내에서 동일한 문자로 표시된 막대는 p에서 크게 다르지 않았습니다. <0.05 Duncan의 다중 테스트에 따른 수준
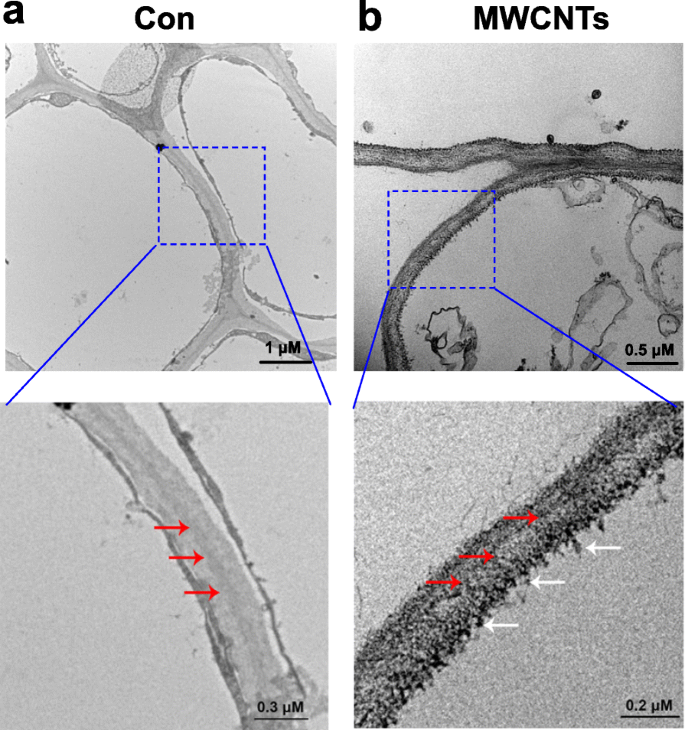
투과전자현미경(TEM)의 도움으로 MWCNT의 분포를 쉽게 평가할 수 있습니다. 그림 3에 나타난 결과는 MWCNT가 외인성으로 적용될 때 토마토 묘목에 흡수될 수 있고 MWCNT의 분포는 뿌리 세포에 있는 것으로 밝혀졌다. 이 결과는 묘뿌리가 MWCNT가 포함된 액체 용액에서 직접 배양되기 때문에 이해할 수 있습니다.
<그림>
토마토 뿌리에서 MWCNT의 분포. 증류수로 처리한 3일 된 토마토 묘목의 TEM 이미지(Con; a ) 또는 5 mg/mL MWCNT(b ) 1 일 동안 복용했습니다. 빨간색 화살표는 세포벽을 나타내고 흰색 화살표는 MWCNT를 나타냅니다.
MWCNT에 의한 NO 합성 및 이후 LR 형성은 NO의 제거제인 cPTIO에 민감했습니다.
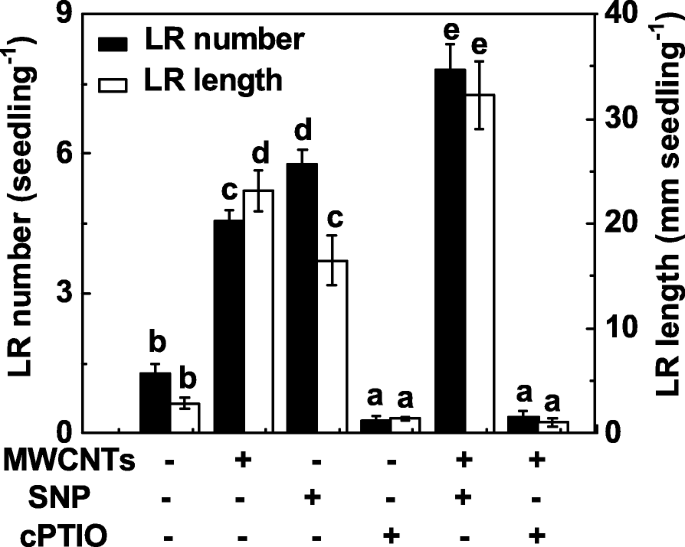
NO가 MWCNT에 의해 유도된 LR 형성에도 관여하는지 여부를 조사하기 위해, MWCNT에 의해 유도된 LR 형성에서 NO의 기능은 NO 방출 화합물과 스캐빈저를 사용하여 내인성 NO 수준을 조작함으로써 평가되었다. 이전 결과와 유사하게[31], Sodium nitroprusside(SNP)의 투여는 LR 형성을 유도할 수 있으며, SNP와 MWCNT를 함께 적용했을 때 부가적인 반응이 관찰되었다(Fig. 4). 2-(4-카르복시페닐)-4,4,5,5-테트라메틸이미다졸린-1-옥실-3-옥사이드 칼륨염(cPTIO; a scavenger of NO)을 첨가했을 때, MWCNT에 의한 LR 형성의 촉진 반응은 유의하게 장애인. 단독으로 cPTIO는 무화학 대조군과 비교하여 LR 발달을 억제할 수 있으며, 이는 뿌리 기관 형성에서 내인성 NO의 중요한 역할을 나타냅니다.
<그림>
MWCNT에 의한 LR 형성은 cPTIO의 제거제인 내인성 NO 제거에 민감했습니다. 3일 된 토마토 묘목을 증류수, 5 mg/mL MWCNT, 0.1 mM SNP, 0.2 mM cPTIO 단독 또는 조합으로 3 일 동안 처리했습니다. 그 후, 유묘당 출아된 측근의 수와 길이(> 1 mm)를 결정하였다. 3개의 생물학적 복제물에 30(10 × 3)개의 식물이 있었고, 실험은 3번 수행되었다. 데이터는 평균 ± SE입니다. 각 실험 세트 내에서 동일한 문자로 표시된 막대는 p에서 크게 다르지 않았습니다. <0.05 Duncan의 다중 테스트에 따른 수준
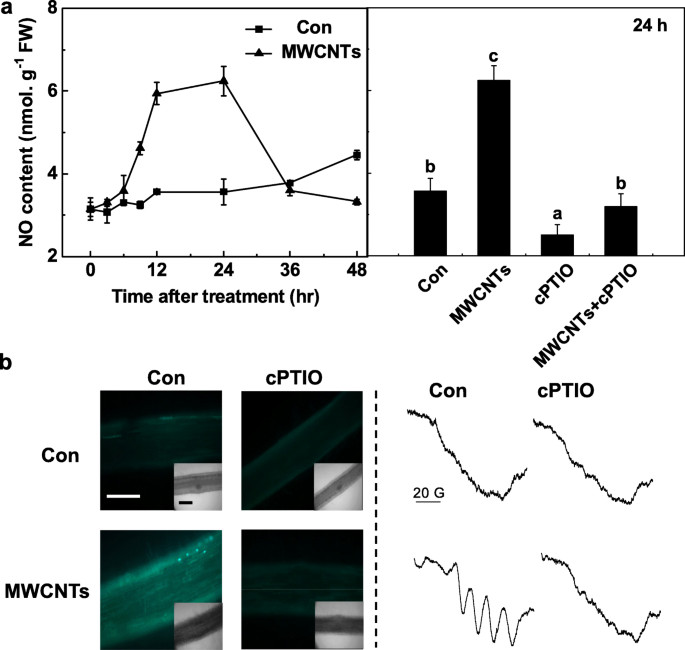
MWCNT 반응에서 내인성 NO의 중요한 역할을 추가로 평가하기 위해, 생체 내 NO 생성의 시간 경과를 먼저 Greiss 시약 방법으로 검출하였다. 위의 결정 과정에서 cPTIO로 전처리된 동일한 여액을 블랭크로 간주하여 정확한 결과를 얻었다. 토마토 묘목 뿌리에서 NO 생산은 MWCNT 처리 후 24 h까지 극적으로 증가하고 초기 수준으로 회복되는 것으로 관찰되었습니다(48 h; 그림 5a). MWCNT에 의해 24시간 동안 촉발된 내인성 NO의 최대 수준 이상은 NO의 제거제인 cPTIO에 의해 분명히 폐지되었으며, 이는 NO의 특정 역할을 시사합니다.
<그림>
MWCNT에 의한 NO 생성은 NO의 제거제인 cPTIO에 의해 차단되었습니다. 3일 된 토마토 묘목은 각각 0.2 mM cPTIO가 있거나 없는 증류수와 5 mg/mL MWCNT로 처리되었습니다. 아 Greiss 시약 방법을 사용하여 결정된 NR 활성(왼쪽) 및 NO 생성(오른쪽)의 변화. ㄴ 24시간 동안 처리한 후 NO 신호는 LSCM(왼쪽)과 EPR(오른쪽)에 의해 분석되었습니다. 스케일 바 =0.1 mm. 데이터는 평균 ± SE입니다. 동일한 문자로 표시된 막대는 p에서 크게 다르지 않았습니다. <0.05 Ducan의 다중 테스트에 따른 수준
위의 결과를 확인하기 위해 LSCM과 ESR을 모두 채택하였다. 첫째, 토마토의 묘 뿌리에서 내인성 NO 수준의 변화는 세포 투과성, 상당히 NO 특이적 형광 프로브 DAF-FM DA를 사용하여 NO를 표지하고 LSCM으로 이미징함으로써 모니터링되었습니다. 이전 결과(그림 5a)와 유사하게 cPTIO의 존재하에서 MWCNT에 의해 촉발된 증가된 DAF-FM 의존 형광 강도는 크게 사라졌습니다(그림 5b). 이러한 결과는 DAF-FM 유발 형광이 토마토 묘목 뿌리의 내인성 NO 수준과 관련이 있음을 시사합니다.
MWCNT에 의한 NO 생성은 EPR 분광법으로 확인하였다. 예상대로, MWCNT로 24시간 동안 처리된 묘목 뿌리는 NO 복합체의 전형적인 초미세 구조 삼중항을 나타냈다. 그러나 cPTIO의 추가는 위의 신호를 폐지하여 MWCNT 노출이 강력한 NO 생성을 초래했음을 나타냅니다(그림 5b). 종합적으로, 이러한 데이터는 토마토 묘목에서 MWCNT 유발 LR 형성에 NO 합성이 필요할 수 있음을 시사했습니다.
NR은 MWCNT로 인한 NO 생성 및 이후 LR 형성에 대한 책임이 있을 수 있음
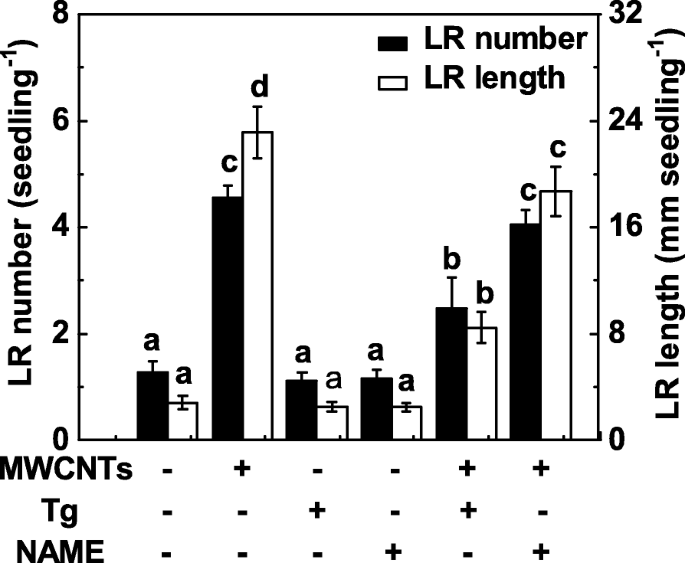
NR 및 포유류 유사 NOS는 식물의 NO 합성과 관련된 두 가지 주요 효소이기 때문에 tungstate(NR 억제제)와 NAME(포유류 NOS 억제제)이 후속 실험에 적용되었습니다. 여기에서, 텅스텐산염 처리는 MWCNT 처리된 토마토 묘목 뿌리에서 LR 형성의 촉진을 실질적으로 억제하였다(그림 6). 상대적으로, MWCNT에 의해 촉발된 LR 형성의 유도는 NAME의 첨가에 의해 강하게 억제되지 않았으며, 이는 포유동물-유사 NOS가 MWCNT에 의해 유도된 NO 생산을 담당하는 표적 NO 합성 효소가 아닐 수 있음을 나타냅니다. 또한 tungstate 또는 NAME을 별도로 적용한 경우 토마토 묘목에서 LR 형성의 약간이지만 유의한 감소가 관찰되지 않는 것으로 관찰되었습니다.
<그림>
MWCNT 및 NO 합성의 두 가지 억제제에 대한 반응으로 LR 형성의 변화. 3일 된 토마토 묘목에 5 mg/mL MWCNT, 20 μM 텅스텐산염(Tg), 0.2 mM NAME을 단독으로 또는 조합하여 3 일 동안 처리했습니다. 그 후, 유묘당 출아된 측근의 수와 길이(> 1 mm)를 결정하였다. 3개의 생물학적 복제물에 30(10 × 3)개의 식물이 있었고, 실험은 3번 수행되었다. 데이터는 평균 ± SE입니다. 각 실험 세트 내에서 동일한 문자로 표시된 막대는 p에서 크게 다르지 않았습니다. <0.05 Duncan의 다중 테스트에 따른 수준
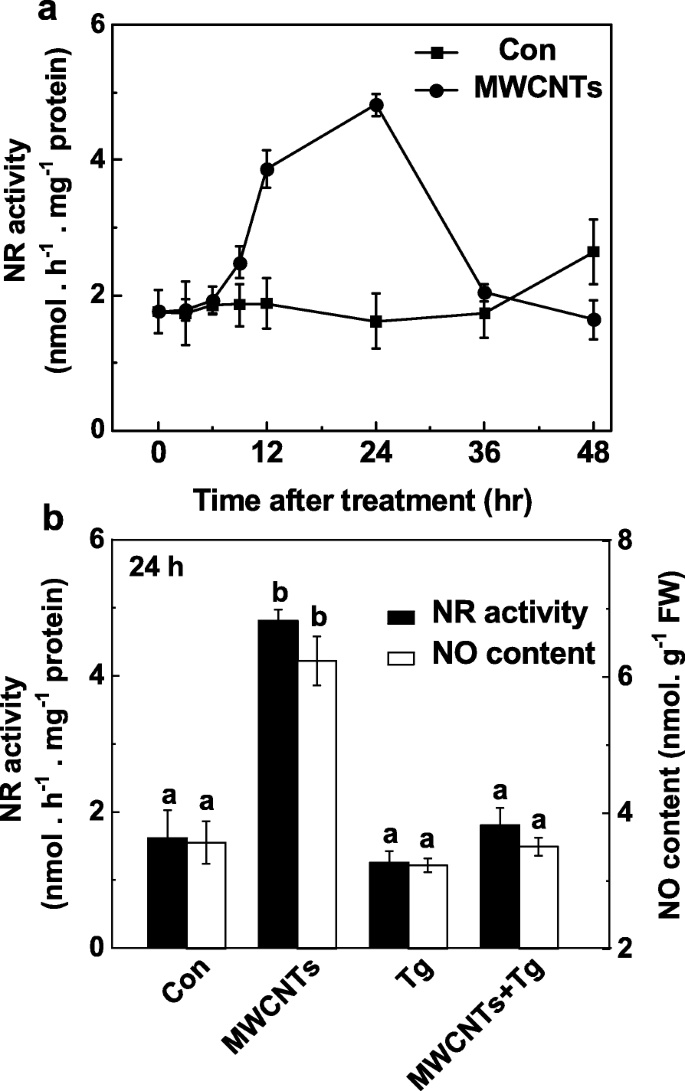
MWCNT에 의해 유발되는 LR 형성에서 NR의 역할은 텅스텐산염이 있거나 없는 적용된 MWCNT에 대한 반응으로 NO 생산을 모니터링함으로써 추가로 조사되었습니다. 내인성 NO 생성의 변화(그림 5a)와 비교하여, NR 활성의 시간 경과 분석은 유사한 경향을 나타내었으며(그림 7a), MWCNT 처리 후 24 h에서도 정점을 이뤘습니다. 이러한 결과는 MWCNT에 의해 유도된 NO 생성의 증가가 주로 NR의 활성 증가로 인한 것일 수 있음을 시사했습니다. 일관되게, 텅스텐산염에 의한 NR 의존적 NO 생성 억제는 Greiss 시약 방법(그림 7b), LSCM 및 EPR(추가 파일 1:그림 S1)을 사용하여 확인되었습니다.
<그림>
MWCNT로 유도된 NO 생성은 NR 억제제인 tungstate에 의해 차단되었습니다. 3일 된 토마토 묘목을 증류수와 20 μM 텅스텐산염(Tg) 유무에 관계없이 5 mg/mL MWCNT로 처리했습니다. NR 활동의 변화(a ) 및 NO 생산(b ) Greiss 시약 방법을 사용하여 결정합니다. 데이터는 평균 ± SE입니다. 동일한 문자로 표시된 막대는 p에서 크게 다르지 않았습니다. <0.05 Ducan의 다중 테스트에 따른 수준
토론
식물 독성은 나노 입자의 잠재적인 환경 영향을 이해하는 데 중요한 고려 사항입니다[4, 7, 61,62,63]. 풍부한 증거는 MWCNT가 산화 손상 유도, 종자 발아, 뿌리 성장 및 발달 억제를 포함하여 식물에 독성이 있음을 보여줍니다[11, 63, 64]. 그러나 SIMR의 표현형으로서 측면 뿌리 형성을 통한 뿌리 분지는 다양한 환경 신호에 대한 뿌리 시스템의 적응성의 중요한 구성 요소입니다[17]. 이 작업에서 우리는 생물학적, 약리학적 및 생화학적 분석을 통합하여 실험 조건에서 적어도 부분적으로 MWCNT 유도 LR 형성에서 NR 매개 NO 생성의 관여를 보여줍니다. 또한 MWCNT에 의해 자극된 뿌리 기관 형성에서 NO의 기능은 식물 발달 과정과 스트레스에 대한 적응에 관여하는 이 두 번째 메신저의 중심 역할을 강조했습니다[29,30,31,32,33, 37, 38].
먼저, 5 mg/mL MWCNT(OD 6–12 nm)가 뿌리 조직으로 들어갈 수 있음을 확인했습니다(그림 3). 그 후, 토마토 LR 형성의 유도가 관찰되었으며(그림 1), 잘 알려진 NO 방출 화합물인 NAA와 SNP(그림 4)의 유도 역할을 모방했습니다[30, 31]. MWCNTs(각각 OD 6–13 nm, 약 9.5 nm 및 30–40 nm)로 공격을 받았을 때 수지 나무[41], 상추[42] 및 Arabidopsis[43]에서 유사한 유도 반응이 발견되었습니다. 예를 들어, 순수한 MWCNT(p-MWCNT) 또는 카르복실 기능화된 MWCNT(c-MWCNT)(평균 직경 9.5 nm)를 사용한 적용은 상추 묘목에서 LR의 발달을 촉진할 수 있습니다[42]. 대조적으로, MWCNT(OD 20-70 nm [40])에 노출되었을 때 대두 식물에서 일차 뿌리 및 심지어 LR 형성의 억제가 동시에 발견되었습니다. MWCNT의 외경 데이터(표 1)와 비교하여 MWCNT가 LR 형성에 미치는 영향은 직경에 따라 다르며 직경이 작을수록 촉진되고 직경이 클수록 억제가 나타난다고 가정했습니다. 물론 관련 메커니즘을 주의 깊게 조사해야 합니다. 식물 염분 내성에서도 유사한 현상이 확인되었다[50]. 이상의 결과와 결합하여 나노물질의 기능은 종에 따라 다를 수 있으며, 그 반대의 경우에도 나노물질의 종류에 따라 다양한 생물학적 효과가 나타날 수 있음을 추론하였다. 그러나 다양한 MWCNT 용량[48] 및 식물 성장 조건과 같은 다른 영향 요인을 쉽게 배제할 수 없습니다.
동일한 농도의 SWCNT, 그래핀 및 AC를 포함한 다른 나노물질과 비교하여 MWCNT에서 LR 형성의 최대 유도 및 싹 성장에 대한 독성 효과가 관찰되었습니다(그림 2). 이는 높은 전기 전도도, 큰 비표면적, 높은 종횡비 및 놀라운 열 안정성을 갖는 나노 물질의 한 유형인 MWCNT의 특수한 물리적 특성과 관련이 있을 수 있습니다[65]. 나노물질의 독성 효과는 오이, 양배추, 당근, 양파 등에서 널리 보고되었다[66, 67].
신호 분자로 작용하는 NO가 환경 적응에서 발달에 이르는 광범위한 식물 과정을 조절할 수 있다는 충분한 증거가 있으며 후자는 종자 발아 및 뿌리 기관 형성을 포함합니다[29, 68,69,70,71,72,73 ]. 우리의 후속 실험은 NO가 MWCNT 유도 LR 형성에 관여할 수 있음을 보여주었습니다. 식물 세포에서 NO 생성을 영상화하는 여러 방법이 적용되었지만 각 방법에는 민감도 부족 및 NO 독립 분자에 의한 간섭과 같은 단점이 존재할 수 있습니다[74]. 따라서, NO의 제거제인 cPTIO의 적용과 함께 Greiss 시약 방법, LSCM 및 EPR을 포함하여 NO 이미징 및 결정을 담당하는 세 가지 방법이 실험 조건에 적용되었습니다. 세 가지 방법을 사용하여 토마토 묘목에서 MWCNT에 의해 유도된 내인성 NO 생성 증가가 NO 제거제인 cPTIO(그림 5)에 의해 제거됨을 관찰했습니다[30,31,32]. 중요하게, 이 과정은 cPTIO가 동시에 적용되었을 때 심하게 차단된 MWCNT 유도 LR 발달의 생물학적 반응과 상관관계가 있었습니다(그림 4).
이러한 반응과 외인성으로 적용된 MWCNT에 의해 유도된 NO의 잠재적 소스에 대한 추가 평가는 NO 생성 및 이후 LR 형성이 NR 활성에 기인할 수 있음을 보여주었습니다. 식물에서 NO 생산은 주로 NR과 포유류 NOS 유사 단백질에서 생성됩니다[28]. 그러나 식물 NOS 유전자는 여전히 확인되지 않았지만[35, 75, 76], 포유류 NOS 효소의 억제제를 사용한 일부 실험에서 NO 생산에서 l-아르기닌 의존성 경로의 증거가 일부 제공되었습니다[36, 76]. NR은 식물에서 NO의 가장 중요한 공급원으로 확인되었습니다[28]. 이전 연구에서는 NR 의존적 NO 생성이 뿌리 구조의 조절에 관여하는 질산염 관련 신호로 기능하는 것으로 나타났습니다[32, 33]. 게다가, NR 의존적 NO 생성은 저온 순응[38], 염분 내성[50] 및 abscisic acid에 의한 기공 폐쇄[77]와 밀접한 관련이 있습니다. 우리의 결과는 tungstate(NR의 억제제)가 특히 LR 길이에서 MWCNT로 유도된 LR 형성을 분명히 손상시켰음을 보여주었습니다(그림 6). 대조적으로, l-NAME(포유류 NOS의 억제제)이 사용되었을 때 LR 길이에서 약간의 감소만 있었고 LR 수에서 유의한 차이가 관찰되지 않았습니다. 일관되게, 생화학적 분석은 NR 활성이 NO 생산의 변화와 병행하여 MWCNT(그림 7a)에 의해 분명히 증가되었음을 보여주었습니다(그림 5a). 위의 응답은 텅스텐산염에 의해 완전히 차단될 수 있습니다(그림 7a, 추가 파일 1:그림 S1). 따라서 우리는 MWCNT에 의해 유도된 내인성 NO 생성 증가가 주로 NR 경로에 기인한다고 추론했습니다. 물론 더 많은 유전적 증거를 조사해야 합니다.
결론
In summary, we provide evidence to show that MWCNT-induced NO production via NR might be required for tomato lateral root formation and this was summarized in Fig. 8. Importantly, above findings provide insights into the intricate molecular mechanism of MWCNTs functions in plants.
Schematic representation of the proposed MWCNT-induced tomato lateral root formation mainly via NR-dependent NO production. The role of mammalian NOS-like enzyme was preliminarily ruled out
데이터 및 자료의 가용성
모든 데이터는 제한 없이 완전히 사용할 수 있습니다.
약어
CO:
Carbon monoxide
cPTIO:
2-(4-Carboxyphenyl)-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide potassium salt