현대 나노기술의 제품인 가공 나노입자(ENP)는 잠재적으로 해양 환경에 영향을 주어 해양 생태계에 심각한 위협이 될 수 있습니다. 그러나 ENP에 대한 해양 식물 플랑크톤의 세포 반응은 아직 잘 확립되어 있지 않습니다. 여기에서는 4가지 서로 다른 규조류 종(Odontella mobiliensis , 그레테 골격종 , Phaeodactylum tricornutum , 탈라시오시라 슈도나나 ) 및 녹조류(Dunaliella tertiolecta) ) 모델 ENP 처리에서 세포외 고분자 물질(EPS) 방출:25nm 이산화티타늄(TiO2 ), 10–20nm 이산화규소(SiO2 ) 및 15–30nm 이산화세륨(CeO2 ). SiO2를 찾았습니다. ENP는 이러한 조류(200–800%)에서 EPS 방출을 크게 자극할 수 있지만 TiO2 ENP 노출은 가장 낮은 방출을 유도했습니다. 또한 세포 내 Ca
2+
의 증가 농도는 ENP에 의해 유발될 수 있으며, 이는 EPS 방출 과정이 Ca
2+
를 통해 매개됨을 시사합니다. 신호 경로. 세포 메커니즘 매개 ENP 유발 EPS 방출에 대한 더 나은 이해를 통해 해양 생태계에 대한 부정적인 영향을 완화하기 위한 잠재적 예방 및 안전 조치를 개발할 수 있습니다.
<섹션 데이터-제목="배경">
배경
1~100nm(적어도 한 치수) 크기 범위의 가공 나노입자(ENP)는 프린터 잉크 및 페인트, 세제, 살균제, 코팅제, 화장품, 자외선 차단제, 타이어, 컴퓨터 구성 및 약물 전달. ENP의 유망한 적용을 감안할 때 미국의 NNI(National Nanotechnology Initiative)에 대한 자금 지원은 2017년에만 14억 달러에 달했습니다[1,2,3]. 나노 규모의 기초 지식을 확립하는 것은 첫 번째 단계에서 나노 기술 연구 커뮤니티의 주요 초점이었습니다. 2009년을 기준으로 이 새로운 지식은 전 세계 시장의 약 4분의 1조 달러를 뒷받침했으며 그 중 약 910억 달러는 나노 스케일 구성 요소를 통합한 미국 제품에 있었습니다[4]. 나노 기술의 급속한 발전으로 ENP가 결국 수중 시스템으로 가는 길을 찾는 것은 불가피합니다. 환경에서 잠재적인 독성(예:활성 산소 종, ROS 생성 가능성) 측면에서 ENP의 주요 관심사는 크고 독특한 표면 반응성과 관련이 있습니다. 그러나 해양 생태계에 미치는 실제 영향은 자연수의 복잡한 환경 및 생물학적 요인과 다양한 ENP로 인해 크게 알려지지 않았습니다[1, 5, 6]. 이전 연구에서는 ENP가 조류 기반 해양 생태계에 심각한 피해를 줄 수 있음을 보여주었습니다[7, 8]. 해양 생물(특히 식물성 플랑크톤)은 ENP와 상호 작용하여 부정적인 영향을 미치는 것으로 나타났습니다[9,10,11]. 다양한 분야에서 나노기술 활용 가능성이 증가함에 따라 점점 더 많은 ENP가 수중 환경으로 유입될 수 있으므로 ENP에 대한 해양 식물성 플랑크톤의 세포 반응은 더 많은 관심을 필요로 합니다[12,13,14,15,16,17,18,19,20, 21].
자가영양 미생물이든 종속영양 미생물이든 대부분의 해양 미생물은 일반적으로 해양 생태계에서 성장 억제제, 성장 촉진제, 독소, 금속 제거제 또는 종속 영양 주기 [22,23,24,25,26]. 해양의 식물성 플랑크톤과 박테리아에서 방출되는 EPS는 다당류가 풍부한 음이온성 콜로이드성 생체 고분자로 해양 겔, 해양 눈, 생물막 형성뿐만 아니라 콜로이드 및 미량 원소 제거 및 다양한 환경 위협으로부터 보호합니다. ENP 포함 [7, 15, 19, 20, 25, 27]. 또한 식물성 플랑크톤이 다양한 스트레스를 겪을 때 EPS의 분비는 자연스러운 반응으로 여겨진다[8].
Ca
2+
다수의 세포내 신호전달 경로에 관여하는 일반적인 2차 메신저입니다. Ca
2+
규조류 Amphora coffeaeformis의 주화성, 운동성 및 접착에 필요합니다. [28]. 강화된 세포내 유리 칼슘
2+
수준은 많은 세포 내 신호 전달 경로에 관여하는 protein kinase C의 활성화를 유도하는 것으로 알려져 있습니다[29]. EPS의 방출은 규조류의 운동성 및 부착과 밀접한 관련이 있기 때문에 Ca
2+
-매개된 분비 과정은 규조류로부터 EPS의 방출을 제어하고 [30] Ca
2+
를 확인하는 직접적인 증거 신호 전달, 세포외 배출 및 Ca
2+
상관 관계 엑소사이토시스를 통한 신호전달은 우리의 이전 연구에서 보고되었습니다[31]. 과거 연구에서도 ENP와의 상호작용이 세포 내 Ca
2+
를 변화시킬 수 있음이 입증되었습니다. 세포 신호 전달에 필수적인 경로 [29, 32,33,34]. 특정 세포내 Ca
2+
농도 변화는 세포 신호 전달 및 분비 과정에서 중요합니다. 그러나 이산화티타늄(TiO2 ), 이산화규소(SiO2 ) 또는 이산화세륨(CeO2 ) 세포내 Ca
2+
변경 식물성 플랑크톤 수준.
2013년에 Quigg et al. [8] 조류에 대한 ENP의 직접 및 간접 독성 효과를 요약했습니다. 이전 실험에서 ENP는 EPS 응집을 촉진하는 것으로 나타났습니다[35]. 이와 관련하여 EPS는 수생 생물에 대한 직접적인 ENP 유발 독성을 악화시키거나 감소시킬 수 있습니다[7, 15, 36]. 그러나 ENP 스트레스 하에서 식물성 플랑크톤으로부터 EPS 방출에 대한 직접적인 측정은 보고된 적이 없습니다. 이 연구의 목적은 4가지 다른 규조류(Odontella mobiliensis , 그레테 골격종 , Phaeodactylum tricornutum , 탈라시오시라 슈도나나 ) 및 녹조류(Dunaliella tertiolecta) ) ENP 처리에서. 식물성 플랑크톤에서 ENP 유발 EPS 방출의 기본 메커니즘을 이해함으로써 예방 및 안전 조치의 구현은 해양 생물에 대한 잠재적으로 해로운 영향을 완화할 수 있습니다.
결과 및 토론
ENP 특성화
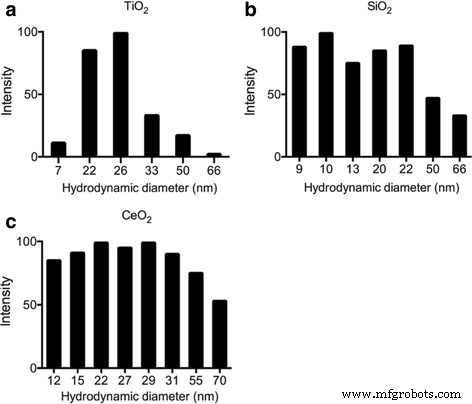
동적 레이저 산란(DLS)을 사용하여 순수한 물에 현탁된 다음 ENP의 크기 메트릭을 특성화했습니다. TiO2 , SiO2 , CEO2 . 입자 크기 분포는 TiO2에서 7~66nm 범위였습니다. , 9 ~ 66nm(SiO2) 및 12~70nm(CeO2) . TiO2의 주된 크기인 반면 일부 더 큰 크기는 응집 또는 덩어리로 인한 것일 수 있습니다. 25nm, SiO2 10~20nm, CeO2 제조업체의 정보와 일치하는 15~30nm입니다(그림 1).
<그림>
a의 DLS 평가에 의한 ENP 특성화 TiO2 , b SiO2 , 및 c CEO2 크기 분포를 보여주는 초음파 처리 후 L1 배지에서. DLS 샘플의 ENP 최종 농도는 1μg/ml이며, 측정 시간은 초음파 처리 직후 3분입니다.
ENP는 세포내 Ca 유도
2+
식물성 플랑크톤의 농도
ENP가 세포 내 Ca
2+
의 증가를 유도할 수 있는지 여부를 조사하기 위해 농도, 식물플랑크톤 세포(OD 600 =0.8)에 Fluo-4AM 염료를 로딩하고 1mg/ml의 25nm TiO2에 노출 , 10–20nm SiO2 및 15–30nm CeO2 ENP 각각. 세포 내 Ca
2+
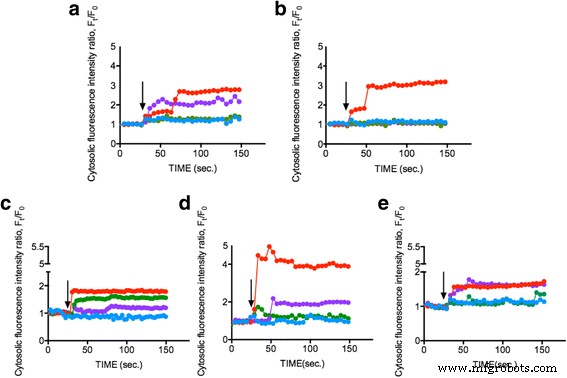
의 변화 식물 플랑크톤 세포 내 형광 강도로 표시되는 농도를 150초 동안 모니터링했습니다. 그림 2a–e는 세 가지 각각의 ENP 중 1mg/ml가 Ca
2+
를 증가시켰음을 보여줍니다. SiO2의 농도 약 50–300%, TiO2 약 40%, CEO2 제어 조건(L1 배지)이 변경되지 않은 상태에서 약 150-200%만큼 감소했습니다. 결과는 ENP가 상당한 세포내 Ca
2+
를 유도할 수 있음을 보여줍니다 식물성 플랑크톤의 반응은 식물성 플랑크톤이 Ca
2+
를 통해 별개의 ENP에 반응한다고 제안합니다. 신호 경로. 우리의 데이터는 세포 내 Ca
2+
의 사소한 변화만을 나타냅니다. TiO2일 때 수준 TiO2로 인한 상당한 식물성 플랑크톤 세포 사멸에 잠재적으로 기인한 존재 -유도 독성 [37, 38]. 이전 연구에서 TiO2 세포 내 Ca
2+
의 증가 촉진 농도 [34]와 상당한 세포 사멸 [39]. 그러나 SiO2 놀랍게도 가장 분명한 세포내 Ca
2+
를 보여주었습니다. 모든 식물성 플랑크톤 종에 대해 증가하는 반면 CeO2 중간 세포내 Ca
2+
만 유발할 수 있습니다. 농도 증가. 이전 연구는 높은 CEO의 잠재력을 시사했습니다2 농도(> 50mg/ml)로 세포 내 산화 스트레스 및 세포 내 Ca 상승 유도
2+
효과는 작았지만 우리의 발견을 뒷받침했습니다[40]. 또한 인공 해수에서 각 ENP의 제타 전위를 측정하여 표면 전하로 인해 발생할 수 있는 잠재적 영향을 해결했습니다. 그러나 값은 낮았다. 측정 결과 ENP가 거의 중립적인 것으로 간주됩니다[41](추가 파일 1:보충 데이터). 이것은 서로 다른 ENP가 세포 내 Ca
2+
를 유도하는 것으로 밝혀진 첫 번째 보고서였습니다. 특정 식물성 플랑크톤의 농도 변화는 궁극적으로 미래 연구를 위한 새로운 길을 열어줍니다.
<사진>
세포내 Ca
2+
측정 다른 ENP에 의한 자극 후 농도. 다양한 식물성 플랑크톤 세포 a두날리엘라 테르티올렉타 , b탈라시오시라 슈도나나 , ㄷ백혈병 , dPhaeodactylum tricornutum , 및 eOdontella mobiliensis TiO2로 처리되었습니다. 25nm(녹색), SiO2 10–20nm(빨간색), CeO2 농도가 1mg/ml인 15~30nm(보라색) 및 대조군(파란색) 검은색 화살표는 EPN이 적용된 시점(30초)을 나타냅니다. 측정값은 평균 20개의 개별 셀에서 얻은 대표적인 데이터를 보여줍니다.
식물성 플랑크톤의 ENP 유발 EPS 방출
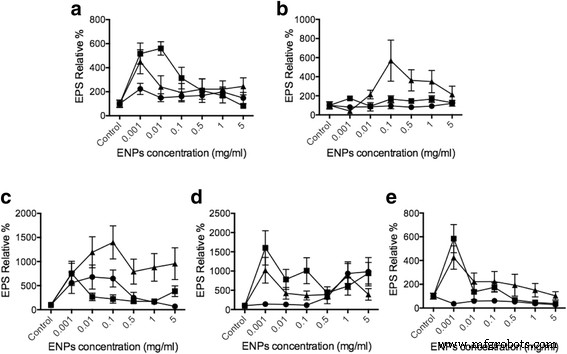
효소 연결 렉틴 분석(ELLA)은 TiO2로 자극되었을 때 식물성 플랑크톤 세포에서 방출되는 EPS의 양을 평가하는 데 사용되었습니다. , SiO2 , CEO2 ENP, TiO2에 대한 이전 연구를 기반으로 하는 농도 범위는 1μg/ml ~ 5mg/ml입니다. [42, 43] 그리고 CEO2 [44,45,46]. EPS 분비는 비교를 위한 동일한 염기를 갖기 위해 총 식물성 플랑크톤 DNA 양(추가 파일 1:보충 데이터)으로 정규화되었습니다. 대조군과 비교하여 10–20nm SiO2Dunaliella에서 EPS 방출을 최대 550%까지 증가시킬 수 있습니다. , 탈라시오시라 500% , 골격종 1000% , Odontella 400% , 900% Phaeodactylum (그림 3). 식물성 플랑크톤 종이 TiO2에 노출되었을 때 , 골격종만 발생하므로 EPS 분비에 대한 강력한 영향은 없었습니다. 및 패오닥틸룸 유의미한 변화를 보였다. 따라서 EPS 방출 데이터는 세포 내 Ca
2+
와 일치합니다. 집중 결과. TiO2 세포내 Ca
+2
농도 변화는 TiO2의 독성으로 인해 매우 제한적인 변화를 보였습니다. 식물성 플랑크톤에. ROS의 생성과 잔류물은 식물성 플랑크톤의 세포 사멸과 같은 많은 합병증을 유발할 수 있습니다[47,48,49]. CEO2에서 치료, 결과는 Dunaliella에서 미미한 효과를 보였습니다. , 골격종 , 오돈텔라 , 및 패오닥틸룸 . 그러나 SiO2Thalassiosira pseudonana에서 가장 유의한 EPS 유도를 나타냈습니다. (약 600%) 및 그레테(Skeletonema grethae) (약 1000-1500%). 이 데이터는 다양한 ENP가 식물성 플랑크톤과 세포 내 Ca
2+
에서 특정 EPS 방출을 유도할 수 있음을 나타냅니다. 변경 사항은 EPS 릴리스 결과와도 일치합니다. 세포 내 Ca
2+
의 변화를 평가함으로써 농도, Ca
2+
에 직접적인 연결이 있음이 분명합니다. ENP가 식물성 플랑크톤에서 EPS 분비를 유발하는 세포 경로. 여기에서의 관찰은 Phaeocystis를 기반으로 한 이전 연구와 일치합니다. EPS 릴리스 [31]. 결과는 식물성 플랑크톤이 Ca
2+
에 의해 조절되는 다양한 EPS 방출에 반응하는 ENP를 감지하고 구별할 수 있다는 직접적인 증거를 제공합니다. 세포 경로.
ELLA를 사용하면 식물성 플랑크톤과 ENP의 상호 작용을 통해 EPS의 방출을 결정할 수 있습니다. 우리의 결과는 식물성 플랑크톤이 SiO2와 상호작용함에 따라 EPS 분비가 크게 증가했음을 나타냅니다. Dunaliella tertiolecta용 , 탈라시오시라 슈도나나 , 및 그레테 골격종 . 이 규조류는 SiO2를 인식하도록 준비된 것으로 보입니다. 입자. 그러나 Phaeodactylum tricornutum , 강한 EPS 분비는 발견되지 않았습니다. 이 차이는 식물성 플랑크톤 종과 ENP 농도에 따라 달라지는 ENP에 의해 유발되는 EPS 방출을 나타냅니다(그림 3). 이전 연구에서 기름 유출은 기름 유출의 부정적인 결과에 대응하기 위해 제안된 대규모 해양 미생물 EPS 방출을 야기했습니다[50]. 또한 Boglaienko와 Tansel은 SiO2 입자는 오일 응집체를 효율적으로 제거할 수 있었습니다[51]. 우리의 발견은 낮은 독성 SiO2 입자는 특정 식물성 플랑크톤에서 EPS 방출을 유도하여 EPS 응집을 촉진하여 잠재적으로 기름 유출 제거를 촉진할 수 있습니다. 이산화세륨은 식물성 플랑크톤 기반 해양 생태계를 교란하는 것으로 보고된 적이 없습니다. 결과는 CEO2를 보여주었습니다. ENP는 Thalassiosira pseudonana를 제외한 모든 식물성 플랑크톤에 영향을 줄 수 있습니다. CEO2 ENP는 SiO2,와 같이 오일 완화 응용 프로그램을 위해 특정 식물성 플랑크톤에서 EPS 방출을 높일 수 있습니다.
결론
ENP-해양 환경 상호 작용은 나노 물질의 현재 및 미래 방출로 인해 점점 더 중요해지고 있습니다. 여기, 우리는 식물성 플랑크톤에 대한 ENP의 주요 효과 중 하나로 향상된 EPS 분비를 보여줍니다. 우리는 또한 다양한 식물성 플랑크톤이 Ca
2+
를 조절함으로써 다양한 ENP 스트레스에 다르게 반응할 수 있다는 증거를 제공합니다. 경로. 그러나 해양 생태계에 대한 ENP의 완전한 평가는 나노 물질과 해양 유기체 간의 상호 작용에 대한 자세한 지식과 이해를 제공하기 위해 추가 조사가 필요합니다.
<그림>
다양한 ENP에 의해 촉발된 EPS 릴리스. 다양한 식물성 플랑크톤 세포 a두날리엘라 테르티올렉타 , b탈라시오시라 슈도나나 , ㄷ백혈병 , dPhaeodactylum tricornutum , 및 eOdontella mobiliensis TiO2로 처리되었습니다. (원), SiO2 (삼각형), CEO2 (사각형), 각각 5mg/ml 및 1mg/ml, 0.5mg/ml, 0.1mg/ml, 10μg/ml, 1μg/ml(n =3)
방법
식물성 플랑크톤 문화
Odontella mobiliensis의 일괄 배양 (CCMP597), 두날리엘라 테르티올렉타 (UTEX999), 그레테 골격종 (CCMP775), Phaeodactylum tricornutum (UTEX646), 탈라시오시라 슈도나나 (Provasoli - Guillard 해양 식물성 플랑크톤 배양 컬렉션, West Boothbay Harbour, MN, USA)는 L1 해양 배지(Sigma, MO, USA)에서 100μmol m
−2에서 14:10(밝은:어두운) 주기로 성장했습니다.
s
−1
무산소 조건에서 24°C. 배양물의 성장 단계는 혈구계산기로 세포 계수를 통해 결정되었습니다.
나노입자 및 특성화
모든 ENP, TiO2 , SiO2 , CEO2 (Sigma-Aldrich, MO, USA)를 사용하기 전에 순수한 물에서 초음파 처리했습니다. ENP는 테스트 전에 여과된 L1 배지(Sigma, MO, USA)로 재구성되었습니다. ENP의 크기는 호모다인 역학 레이저 산란(DLS)을 사용하여 독립적으로 확인되었습니다. 간단히 말해서, 해수 샘플을 0.22μm Millipore 멤브레인(0.1N HCl으로 사전 세척)을 통해 재여과하고 5개의 10ml 산란 셀에 직접 부은 다음 Brookhaven BI-200SM 레이저 분광계(Brookhaven Instruments, 뉴욕, 미국). 45°각도에서 검출된 산란강도 변동의 자기상관함수는 Brookhaven BI 9000ATautocorrelator에 의해 온라인으로 처리되었고, 입자크기 분포는 CONTIN 방법(Provencher, 1982)에 의해 계산되었다. 각 샘플의 결과는 초음파 처리 직후에 3중으로 수집되었습니다. DLS 분광계의 보정은 단분산 라텍스 미소구체의 표준 현탁액을 사용하여 수행되었습니다(Polysciences, PA, USA).
ENP 치료
식물성 플랑크톤 세포를 L1 배지가 포함된 96웰 플레이트에서 24시간 동안 배양했습니다. 세포를 ENP 스톡으로 처리했습니다:5mg/ml 및 1mg/ml, 0.5mg/ml, 0.1mg/ml, 10μg/ml, 1μg/ml의 TiO2 , SiO2 , CEO2 (Sigma-Aldrich, MO, USA) 또는 48시간 동안 L1 배지(대조군) 분비된 EPS를 포함하는 상층액을 수집하고 4000rpm에서 잠시 원심분리하여 잔류 ENP를 제거했습니다. 이 프로토콜은 이전 간행물[34]에서 수정되었습니다. 여기에 사용된 농도 범위는 환경에서 현재 ENP 수준을 나타내거나 모방하기 위한 것이 아니라 해양 식물 플랑크톤에 대한 ENP의 전체 잠재적 영향을 평가하고 관련 세포 메커니즘을 조사하는 것을 목표로 합니다. 유망한 신생 나노 물질로서 ENP는 아직 완전한 상업 능력에 도달하지 못했습니다. ENP가 바다에 더 많은 ENP를 도입하기 위해 상업 및 가정용 제품 시장에 진입하기 전에 완전한 생태학적 영향에 대한 자세한 평가가 훨씬 필요합니다.
효소 결합 렉틴 분석(ELLA)
분비된 다당류를 포함하는 상층액을 수집하고 1700 rcf(Megafuge 1.0R)에서 간단히 원심분리하여 잔류 ENP를 제거했습니다. 그런 다음 상층액을 4°C에서 밤새 96웰(Nunc MaxiSorp, VWR, CA, USA) 플레이트에서 인큐베이션했습니다. 그 후 96웰 플레이트를 PBST(PBS + 0.05% Tween-20) 및 PBS로 세척한 다음 1% BSA로 차단했습니다. 96웰 플레이트를 PBST 및 PBS로 다시 세척하고 렉틴(Concanavalin A, ConA)(Sigma-Aldrich, MO, USA)과 함께 배양하고 양고추냉이 퍼옥시다제(HRP; 5mg/ml)(Sigma-Aldrich, MO)에 접합했습니다. , 미국), 37°C에서 1시간 기질인 3,39,5,59-테트라메틸벤지딘(TMB; Sigma-Aldrich, MO, USA)을 실온에서 각 웰에 첨가한 후 H2를 첨가하였다. 반응을 종료하기 위해 SO4(Sigma-Aldrich, MO, USA). 광학 밀도는 PerkinElmer VICTOR3(MA, USA)에 의해 450nm에서 측정되었습니다. 이 프로토콜은 이전 간행물[34, 52]에서 수정되었습니다.
DNA 결정
식물성 플랑크톤을 함유한 펠렛을 수집하여 ZR-96 Quick-gDNA 키트(ZYMO Research, CA, USA)를 얻었다. 간단히 말해서, 4x 용해 완충액을 사용하여 식물성 플랑크톤 세포를 부수고 DNA 결합 컬럼을 통해 흐르게 했으며, 결국 용출 완충액에 의해 용리되었습니다. DNA 농도는 NanoDrop ND-1000(Thermo, CA, USA)으로 측정했습니다. 프로토콜은 제조된 키트 프로토콜에서 채택되었습니다.
세포내 Ca 측정
2+
ENP에 의해 유도된 농도
그런 다음 식물성 플랑크톤 세포에 Fluo-4AM 염료(1mM)(Kd =335nM, λEx =494nm, λEm =506nm, ThermoFisher, CA, USA)를 60분 동안 로딩했습니다[31]. 염료 로딩 후 식물성 플랑크톤 세포를 헹구고 L1 배지와 함께 배양하고 1mg/ml TiO2로 처리했습니다. , SiO2 , CEO2 각기. 모든 칼슘 신호 전달 실험은 Nikon 현미경(Nikon Eclipse TE2000-U, Tokyo, Japan)에서 수행되었습니다. 프로토콜 및 조건은 이전 간행물 [31, 34]에서 조정되었습니다.
ENP 측정의 제타 전위
ENP의 표면 전하를 측정하기 위해 ENP의 제타 전위(ζ)는 25°C의 인공 해수가 존재하는 Malvern의 Zetasizer Nano ZS로 측정되었습니다. 각 샘플에서 데이터를 수집한 후 기록된 값의 평균을 냈습니다.
통계 분석
데이터는 평균 ± SD로 보고됩니다. 각 실험은 최소 3회 독립적으로 수행되었습니다. 히스토그램은 GraphPad Prism 6.0으로 만들었습니다. (GraphPad Software, Inc., San Diego, CA, USA).