금 나노로드 코어/은 쉘 나노구조에 의해 유도된 유전독성에서 ROS 및 세포 주기 정지의 역할
초록
은 나노입자(AgNPs)와 은 이온에 의해 간에서 유도된 유전독성을 이해하기 위해 조작된 금 나노막대 코어/은 쉘 나노구조(Au@Ag NR) 및 인간화된 간세포 HepaRG 세포가 이 연구에서 사용되었습니다. 0.4–20 µg mL
−1
에 의해 유도된 DNA 및 염색체 손상의 산화 스트레스 및 세포 주기 정지 관련 Au@Ag NR은 Comet assay, γ-H2AX assay, 소핵시험으로 조사하였다. 또한, Au@Ag NR의 분포를 분석하였다. 우리의 결과는 Ag
+
Au@Ag NR은 HepaRG 세포에서 DNA 절단 및 염색체 손상(염색체 형성)을 일으키고 핵에 보유된 Au@Ag NR은 Ag
+
를 추가로 방출할 수 있습니다. , 주로 세포주기 정지 및 ROS 형성으로 인한 손상을 악화시킵니다. 결과는 세포내 축적, Ag
+
사이의 상관관계를 보여줍니다. 이온 방출 및 AgNP의 잠재적 유전독성.
소개
크기가 1~100nm인 은 나노입자(AgNPs)는 병원체에 침투하고 대사 효소의 내부 설프히드릴 그룹을 비활성화함으로써 광범위한 항균 특성을 나타낼 수 있습니다[1]. 그들은 Escherichia Coli에서 강력한 정균 및 살균 효과를 입증했습니다. , 임균(Neisseria gonorrhoeae) 및 클라미디아 트라코마티스 , 의료용 코팅, 가정용품[2], 상처 드레싱[3]으로 널리 사용됩니다. 강력한 증거는 나노 입자가 핵에 들어갈 수 있고 DNA의 합성 및 전사 과정을 방해할 수 있음을 보여줍니다[4]. 이전 연구에서 우리는 5mg/kg AgNPs의 단일 정맥 투여가 Sprague-Dawley 쥐의 골수 세포에서 현저한 염색체 파손을 유발할 수 있다고 보고했습니다[5]. 10mg/kg 이상의 AgNP를 복강 내 단일 주사하면 DNA와 염색체 손상이 모두 유발됩니다[6]. Flower et al. [7]은 AgNP가 50 및 100μg mL
−1
용량에서 투여 후 5분 이내에 DNA 손상을 유발할 수 있으며, 빠르게 방출되는 은(Ag)의 유전독성을 강조합니다. 과도한 노출의 위험성을 고려하여 나노유전독성학이나 조작된 나노물질의 DNA 손상 및 발암 가능성에 대한 연구가 많은 주목을 받고 있다[8].
AgNP에 의한 유전적 손상의 주요 기전은 반응성 산화 종의 과잉 생산, 염증 및 세포주기 교란으로 간주됩니다 [9, 10]. 이전 연구에서 제안된 바와 같이 AgNPs는 산화적 손상을 통해 DNA와 직접 상호작용할 수 있고[11], DNA 수준에서 간기 및 염색체 수준에서 유사분열을 방해하거나, 핵단백질 및 유사분열 방추 장치와 상호작용하여 세포 주기 체크포인트를 방해할 수 있습니다[11]. 12]. 그러나 AgNPs에 의해 유도된 유전독성이 부분적으로 나노입자에 기인하는지 [13, 14] 아니면 완전히 방출된 Ag
+
이온은 여전히 불분명합니다[15, 16].
AgNPs의 유전 독성 조사는 조직에서 은의 불안정하고 중단되지 않은 방출로 인해 어렵습니다. 이는 AgNPs의 위치를 파악하고 나노 코어를 Ag와 구별하는 데 어려움을 초래합니다. 우리 그룹은 최근 나노입자에 의해 유도된 독성을 연구하기 위해 금 나노로드 코어/은 쉘(Au@Ag NR) 나노구조를 개발했습니다[17]. Au@Ag NR의 금핵은 조직에 생리학적으로 타고난 것이며 Ag
+
의 방출을 모니터링하기 위한 내부 표준으로 사용될 수 있습니다. 유도 결합 플라즈마 질량 분석기(ICP-MS)를 사용하여 측정된 Ag/Au 비율의 변화를 모니터링하여 막대에서 이온을 제거합니다[18]. 이 방법으로 독성의 다양한 기원을 확인할 수 있습니다. 이전 연구에 따르면 방출된 Ag
+
Au@Ag NR 껍질의 이온은 신장 산화 손상을 일으키고 결국 형태학적 변화와 사구체의 여과 기능 손상을 초래합니다[19]. Jiang et al. [20]은 Au@Ag NR에 의한 입자 특이적 활성과 세포 내 은 이온 방출이 모두 과립막 세포의 독성 반응에 기여한다고 제안했습니다. 우리는 또한 Au@Ag NR을 AgNPs의 생체 내 유전독성 가능성을 연구하기 위한 모델로 채택했으며 돌연변이 유발성이 아닌 세포 분열이 Ag 껍질과 방출된 Ag
+ 독성 패턴에는 차이가 없었지만 [21].
간은 AgNPs가 축적되기 쉬운 주요 장기 중 하나이며 AgNPs에 의한 유전독성의 표적 장기/조직으로 인식되고 있습니다. 우리의 이전 연구는 Au@Ag NR 1회 정맥 투여 후 8주 동안 SD 쥐의 간에 일정량의 은(8.26 ± 3.90μg/g)과 금(80.07 ± 64.72μg/g)이 남아 있음을 보여주었습니다. ]. 본 연구에서는 인간 간암 유래 HepaRG 세포에서 Au@Ag NR을 이용하여 AgNP로 유도된 염색체 및 DNA 손상에 대한 세포 주기 정지 및 반응성 산화 스트레스의 역할을 규명하고자 하였다. Comet assay, γ-H2AX assay 및 micronucleus test를 포함한 유전독성 시험은 DNA/염색체 손상에서 활성산소(ROS)의 기여도를 조사하기 위해 산화 라디칼 제거제와 병행하여 수행되었으며, 세포 사멸, 세포 주기 및 관련 단백질 AgNPs가 DNA의 합성과 복제를 방해하는 메커니즘을 탐구하기 위해 결정되었습니다. 또한 Au@Ag NR의 세포 내 축적 및 분포는 유도 결합 플라즈마 질량 분석기(ICP-MS)와 투과 전자 현미경(TEM)을 결합하여 나노 입자 및 방출된 Ag 이온의 역할을 구별하여 조사했습니다.
자료 및 방법
세포 배양 및 치료
인간 간암 세포주 HepaRG(Thermo Fisher Scientific)가 이 연구에 사용되었습니다. 세포는 5% CO2의 습한 분위기에서 10% 소태아혈청(FBS, Australia Origin, Gibco) 및 1% 페니실린-스트렙토마이신-글루타민 용액(Gibco)을 포함하는 RPMI 1640에서 배양되었습니다. 37°C에서 세포는 각각 24시간 또는 72시간 동안 증가하는 농도의 Au@Ag NR로 처리되었으며 농도는 IC50에 따라 결정되었습니다. 세포 생존력 분석에 의해 추정. 유전독성에서 ROS의 역할을 조사하기 위해 1mM N -Acetyl-l-cysteine(NAC, Sigma-Aldrich)을 Au@Ag NR로 처리하기 1시간 전에 적용했습니다.
ATP 세포 성장/생존 분석
세포를 96웰 플레이트에 5 × 10
3
의 밀도로 시딩했습니다. /잘. 24시간 배양 후 배지를 흡인하고 세포를 각각 24시간 또는 48시간 동안 다양한 농도의 Au@Ag NR에 노출시켰습니다. 광범위한 농도 스펙트럼이 준비되었으며 처리당 4개의 웰이 한 처리 기간에 수행되었습니다. Au@Ag NR의 세포독성은 생존 세포의 중요한 대사 매개변수인 ATP의 양을 정량화하여 세포 대사 활성을 측정하는 ATP(아데노신 삼인산) 분석(CellTiter-Glo® 2.0 Assay, Promega)으로 조사했습니다. VICTOR Multilabel Plate Reader(2030-0050, PerkinElmer) 및 IC50을 사용하여 생존 세포의 양을 반영하는 발광 신호를 검출했습니다. 값은 Prism 7(GraphPad Prism 7, CA, USA)에 의한 최대 생존율의 절반에 대한 Au@Ag NR의 농도로 추정되었습니다. 생존율은 다음 방정식을 사용하여 계산됩니다.
여기서 RLU는 4개의 우물, RLUvehicle의 평균값으로 표시되는 상대 조명 단위입니다. 나노로드로 처리되지 않은 세포 및 RLU샘플 Au@Ag NR의 다른 농도로 처리된 세포를 나타냅니다.
세포 내 은과 금의 농도 측정
세포 샘플은 마이크로파 분해 시스템을 사용하여 질산에서 분해되었습니다. 소화 후, 샘플은 1% 질산과 염산을 포함하는 혼합물로 준비되었습니다. 용액에서 Ag 및 Au의 양은 ICP-MS(NexION300X, PerkinElmer)에 의해 결정되었습니다. TEM 분석을 사용하여 세포에서 Au NR 및 Au@Ag NR의 존재를 확인했습니다. 세포 샘플을 2.5% 글루타르알데히드와 2% 파라포름알데히드의 혼합물에 4°C에서 2시간 동안 고정했습니다. 세포 펠릿을 고정하고 인산 완충액(pH 7.4)에서 3회 헹구고 4°C에서 2시간 동안 1% 사산화 오스뮴에 후고정했습니다. 이후 샘플을 증류수로 3회 헹구고 15분 동안 서로 다른 농도의 에탄올(각각 50%, 70%, 90% 및 100% 에탄올)에서 차례로 탈수했습니다. 그 후, 1:1 및 1:3 희석의 프로필렌 옥사이드를 20~26°C에서 2시간 동안 수지에 적용했습니다. 중합은 35°C에서 16시간, 45°C에서 8시간, 55°C에서 14시간, 65°C에서 48시간 동안 단계적으로 가열하여 수행했습니다. 초박막 절편을 25분 동안 uranyl acetate와 lead citrate로 염색하고 투과전자현미경(H-7650, HITACHI, Japan)으로 분석했습니다.
기존 및 수정된 혜성 분석
세포를 12웰 플레이트에 2 × 10
5
의 밀도로 시딩했습니다. /웰 또는 3 × 10
5
/well은 각각 24시간 또는 72시간 치료에 사용됩니다. 과산화수소(H2 O2 ) 200μmol의 농도에서 양성 대조군으로 세포에 1시간 동안 노출되었습니다. 각 샘플에 대해 기존 처리 및 Fpg(formamidopyrimidine glycosylase) 처리를 위해 두 개의 웰을 준비했습니다. 기존의 혜성 분석은 이전에 설명한 대로 알칼리성 조건(pH> 13)에서 수행되었습니다[21]. Fpg 처리된 웰의 경우 DNA 풀기 절차 전에 추가 Fpg 처리를 적용하고 슬라이드를 효소 완충액(0.1M KCl, 0.5mM EDTA, 40mM HEPES, 0.2mg.mL
-1
BSA) 각 5분씩 3회 Fpg(New England Biolabs, Inc., UK)를 효소 완충액으로 1:50,000으로 희석했습니다. 희석된 효소의 100밀리리터 분취량을 현미경 슬라이드의 각 겔에 첨가하고 37°C의 습도 챔버에서 30분 동안 인큐베이션했습니다. 나머지 단계는 기존 처리와 동일하였다. 혜성 분석은 삼중으로 수행되었습니다. 샘플당 최소 50개 세포는 Nikon Eclipse 80i 형광 현미경(Nikon, Tokyo, Japan)을 사용하여 독립적으로 점수를 매겼고 Komet 6.0(Andor Technology, Belfast, UK)을 사용하여 꼬리와 올리브 꼬리에서 DNA 백분율의 중간 값을 분석했습니다. 각 샘플의 모멘트(OTM).
유세포 분석 및 고함량 스크리닝에 의한 γ-H2AX 병소의 자격
유세포 분석을 사용한 정량화를 위해 세포를 12웰 플레이트에 2 × 10
5
의 밀도로 시딩했습니다. /웰 또는 3 × 10
5
/well은 각각 24시간 또는 72시간 처리, 고함량 스크리닝 분석의 경우 세포를 6 × 10
3
밀도로 96웰 플레이트에 접종했습니다. /웰 또는 1 × 10
4
/well은 각각 24시간 또는 72시간 치료에 사용됩니다. 양성 대조군으로 2μM 메틸 메탄설포네이트(MMS, Sigma-Aldrich)를 1시간 동안 세포와 병행하여 적용했습니다. 세포를 트리스 완충 식염수(TBS)로 헹구고 실온에서 15분 동안 4% 파라포름알데히드로 고정했습니다. TBS로 세척한 후 세포를 - 20°C에서 30분 동안 50μL 얼음처럼 차가운 메탄올과 함께 배양했습니다. 세포를 TBS에서 3회 더 헹구고 차단 시약(0.3% Triton X-100 및 10% 염소 혈청을 포함하는 TBS)을 1시간 동안 적용했습니다. 1차 항체(마우스 항-포스포-H2AX Ser139, Millipore)를 차단 시약으로 1:200으로 희석하고 4°C에서 밤새 세포와 함께 인큐베이션했습니다. 그런 다음 플레이트를 다시 TBS로 3회 헹구고 차단 시약으로 1:20 비율로 희석한 이차 항체(Alexa Fluor 488 염소 항마우스, Life Technologies)를 추가했습니다. 샘플을 실온에서 1시간 동안 암실에 보관하고 2μg mL
−1
(20μL/웰) DAPI(Invitrogen)를 각 웰에 첨가했습니다. 형광은 유세포 분석기(FACSCalibur, BD Bioscience, NJ, USA) 또는 High Content Analysis System(Operetta CLS, PerkinElmer)을 사용하여 측정되었습니다. 유세포 분석법을 위해 그룹당 최소 10,000개 세포의 데이터를 분석하고 실험을 세 번 수행했습니다. 고함량 분석을 위해 각 웰의 20개 시야와 각 그룹의 최소 5개 웰을 분석했습니다.
세포질 분열 차단 소핵 세포분열(CBMN-cyt) 분석
CBMN-cyt는 Fenech et al.에 의해 설명된 절차에 따라 수행되었습니다. [22]. 세포를 12웰 플레이트에 2 × 10
5
의 밀도로 시딩했습니다. /웰 또는 3 × 10
5
/well은 각각 24시간 또는 72시간 치료에 사용됩니다. 0.2μg mL
−1
Mitomycin C(MMC, Tokyo Chemical Industry Co., Ltd. Japan)는 24시간 동안 양성 대조군으로 세포에 노출되었습니다. 3μg mL
−1
24시간 또는 72시간 처리 후 사이토칼라신 B를 적용하여 세포질 분열 과정을 차단하고 40시간 후에 세포를 수확했습니다. 샘플은 예열된 0.075mol L
−1
로 저장성 후 5% Giemsa로 염색되었습니다. 메탄올과 아세트산의 3:1 혼합물로 KCl 및 고정. 그룹당 3중 웰을 준비하고 웰당 최소 1000개의 이핵 세포를 검사했습니다.
MDA, 총 GSH 및 SOD 함량 측정
세포는 5 × 10
5
의 밀도로 12웰 플레이트에서 배양되었습니다. /웰 또는 3 × 10
5
/well은 각각 24시간 또는 72시간 치료에 사용됩니다. 이어서, 세포를 수확하고 인산완충식염수(PBS)로 3회 헹구었다. 세포 균질물에서 말론디알데히드(MDA)의 양은 티오바르비투르산 기반 방법(Nanjing Jiancheng Bio-engineering Institute, Nanjing, China)을 사용하여 결정되었습니다. 총 글루타티온(GSH) 및 슈퍼옥사이드 디스뮤타제(SOD)의 양은 총 글루타티온 정량화 및 SOD 분석 키트(Dojindo Molecular Technologies, Inc. Kumamoto, Japan)를 사용하여 각각 결정되었습니다. 각 웰의 광학 밀도(O.D)는 VICTOR Multilabel Plate Reader(2030-0050, PerkinElmer)를 사용하여 측정되었습니다.
세포 주기에 대한 유세포 분석
세포는 1 × 10
6
의 밀도로 6웰 플레이트에서 배양되었습니다. /웰 또는 5 × 10
5
/well을 각각 24시간 또는 72시간 처리한 다음 4°C에서 밤새 70% 에탄올로 고정했습니다. 샘플을 PBS로 3회 헹구고 실온에서 15분 동안 PI/Rnase 염색 완충액(BD Biosciences)으로 염색했습니다. 20,000개 세포 중 G0/G1, S 및 G2/M기 아래의 세포 집단은 FL2 면적 대 FL2 너비를 갖는 영역을 사용하여 결정되었습니다. 분석은 유세포분석기(FACSCalibur, BD Bioscience, NJ, USA) 및 FlowJo(BD Bioscience)로 수행되었으며 실험은 3회 수행되었습니다.
세포 사멸의 유세포 분석
세포는 1 × 10
6
의 밀도로 6웰 플레이트에서 배양되었습니다. /웰 또는 5 × 10
5
/well은 각각 24시간 또는 72시간 치료에 사용됩니다. 이후 PBS로 두 번 헹구고 500μL 1 × 결합 완충액(FITC Annexin V Apoptosis Detection Kit I, BD Bioscience)으로 희석하여 현탁액을 약 1 × 10
6
으로 조정했습니다. 세포/mL, 후속적으로 100μL 희석액을 5μL FITC Annexin V 및 5μL PI와 혼합했습니다. 샘플을 실온에서 15분 동안 염색하고 유세포 분석법(FACSCalibur, BD Bioscience, NJ, USA) 및 FlowJo를 사용하여 FL1H 대 FL2H가 있는 영역을 사용하여 초기 및 후기 세포자멸사에서 세포 집단을 결정하기 위해 최소 10,000개의 세포를 분석했습니다. (BD 바이오사이언스). 실험은 세 번 수행되었습니다.
서부 얼룩 분석
세포는 75cm
2
1 × 10
7
밀도의 플라스크 /웰 및 6 × 10
6
/well은 각각 24시간 및 72시간 치료에 사용됩니다. 프로테아제 억제제(PMSF)를 함유하는 RIPA 용해 완충액으로 세포를 용해시키고, BCA 단백질 정량 키트(Beyotime Biotechnology, China)를 사용하여 단백질 농도를 측정하였다. 샘플의 농도는 변성 전에 95°C에서 3분 동안 가열하여 RIPA 용해 완충액을 사용하여 조정했습니다. 단백질 샘플을 12% SDS 폴리아크릴아미드 겔에서 전기영동으로 분리하고 니트로셀룰로오스 막(Millipore)으로 옮겼습니다. 막을 5% 탈지유로 30분 동안 차단하고 1차 p53(SC-137174, Santa Cruz), p21(SC-6246, Santa Cruz) 및 β-액틴(sc-47778, Santa Cruz) 및 2차 항체와 함께 배양했습니다. 염소 항-마우스 IgG(H+L)-HRP(SE131, solabio), 각각. ECL(Enhanced chemiluminescence) 방법을 사용하여 샘플에서 표적 단백질의 발현 수준을 시각화하고 ImageJ 시스템(National Institutes of Health)으로 분석했습니다.
통계 분석
데이터는 평균 ± SEM으로 표시되었습니다. 일원 분산 분석(ANOVA)을 사용하여 음성 대조군과 치료 그룹 간의 차이의 통계적 유의성을 테스트한 다음 SPSS(버전 22, IBM, Armonk, NY, USA)를 사용한 Dunnett 다중 비교 테스트를 수행하고 데이터를 고려했습니다. P에서 통계적으로 유의미함 <0.05. 수치는 Windows용 GraphPad Prism 7(GraphPad Software, La Jolla, CA, USA)을 사용하여 작성되었습니다.
<섹션 데이터-제목="결과">
결과
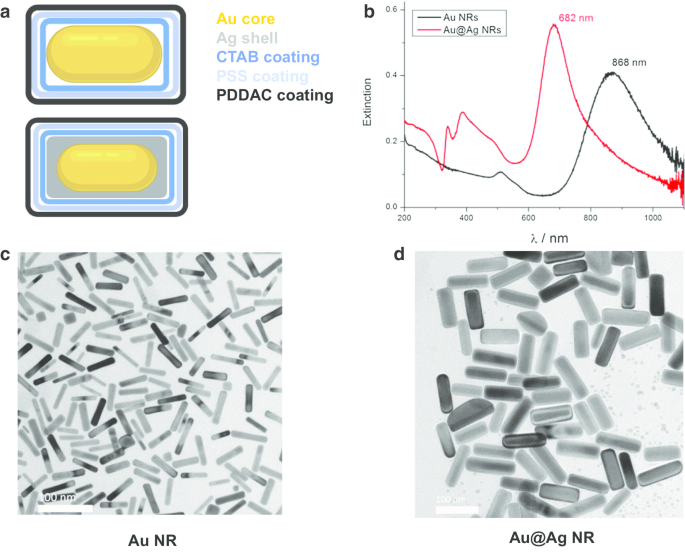
Au NR 및 Au@Ag NR의 특성
금 나노로드(Au NRs), 금 나노로드 코어 및 은 쉘 나노구조(Au@Ag NR)는 이전에 설명된 대로 설계, 준비 및 특성화되었습니다[21]. 간단히 말해서, 평균 직경과 길이는 Au NR의 경우 15.0 ± 2.5nm, 66.7 ± 2.5nm이고 Au.@Ag NR의 경우 26.2 ± 3.0nm, 72.7 ± 8.9nm입니다. Ag 셸 두께는 약 5nm입니다. 물에 분산된 PDDAC 코팅된 Au NR 및 Au@Ag NR의 제타 전위는 각각 37.7 ± 1.6mV 및 52.5 ± 1.4mV였습니다. 준비된 Au@Ag NR의 Ag/Au 중량비는 2.3으로 추정되었다. 특성화 결과는 그림 1에 나와 있습니다.
<그림>
Au NR 및 Au@Ag NR의 특성화. 아 Au NR 및 Au@Ag NR의 구조도; ㄴ 물에 분산된 Au NR 및 Au@Ag NR의 UV-Vis-NIR 소멸 스펙트럼; ㄷ Au NR의 대표적인 TEM 이미지; d Au @Ag NR의 대표적인 TEM 이미지
세포 생존율
Au@Ag NR의 HepaRG 세포에 대한 세포독성은 ATP 생존력 분석(표 1)에 의해 조사되었으며, 세포는 0.125~160μg mL
-1의 다양한 농도에서 Au@Ag NR에 24시간 또는 48시간 동안 노출되었습니다.
. Au@Ag NR은 24시간 및 48시간 노출 후 시간 및 용량 의존적 방식으로 상당한 세포독성 효과를 유도했으며, % 생존력 IC50 20µg mL
−1
에서 및 6µg mL
−1
, 각각 소프트웨어 GraphPad Prism 7.0에 의해 적합합니다. 전반적인 세포 독성을 고려하여 처리 기간을 24시간 및 72시간으로 조정하고 적용 농도는 0.8 µg mL
-1
로 결정했습니다. , 4 µg mL
−1
및 20µg mL
−1
. 또한 Au NR은 불활성 대조군으로 포함되었으며 AuNR 그룹의 Au 함량은 20 µg mL
−1
와 동일했습니다. Au@Ag NR, 16µg mL
−1
. 대조적으로, 산화 스트레스 반응에 대한 대조군으로 Au@Ag NR + NAC 그룹에서 1 mM NAC 전처리가 채택되었습니다(Au@Ag NR의 농도는 20 µg mL
−1
).
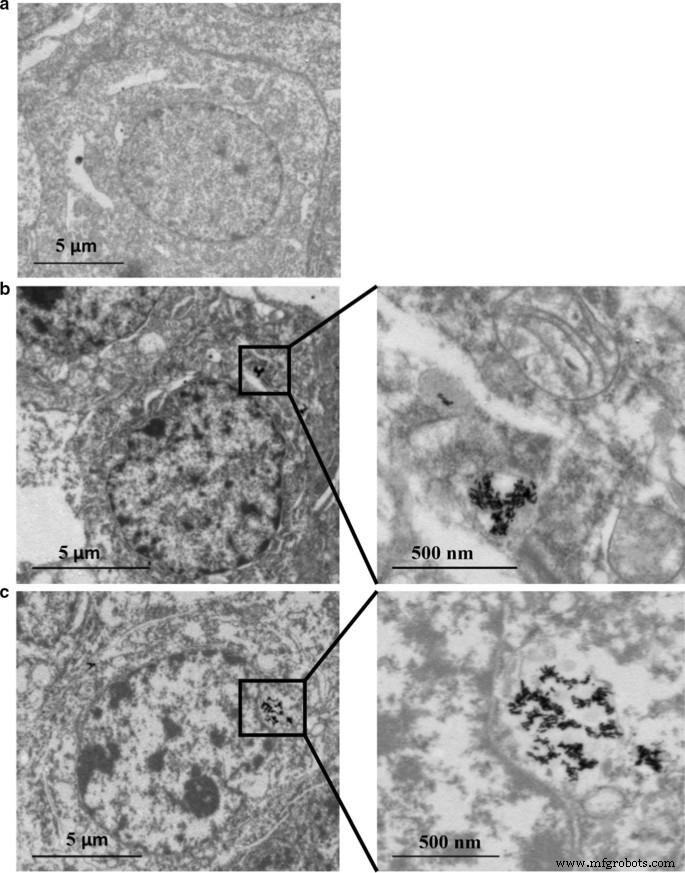
Au NR 및 Au@Ag NR의 세포 분포
HepaRG 세포의 Au 및 Ag 함량 분포를 ICP-MS로 분석했습니다. 표 2 및 3에 나타난 바와 같이 Ag 함량은 용량 의존적으로 증가하였다. 그러나 항산화제 N - Au@Ag NR의 동일한 농도(20 µg mL
−1
)에도 더 적은 Ag 함량이 관찰되었기 때문에 자유 라디칼 제거제로서의 아세틸-l-시스테인(NAC)은 나노입자의 세포 흡수를 제한할 수 있습니다. ) 이 그룹에 적용되었습니다. 24시간에서 72시간으로 Ag/Au 비율의 감소는 Ag
+
의 지속적인 방출을 나타냅니다. Au@Ag NR의 껍질에서. 또한 Ag의 세포 흡수는 Au보다 훨씬 많습니다(표 4). 또한, TEM 데이터는 대부분의 Au NR 및 Au@Ag NR이 덩어리로서 세포에 유지되었음을 보여주었다. Au NR 또는 Au@Ag NR이 핵으로 들어가지 않고 노출된 세포 내부에서 나노로드의 구조가 명확하게 보였다(그림 2).
<그림>
Au NR 및 Au@Ag NR 내재화:16μg mL에 24시간 노출 후 80kV에서 TEM에 의한 HepaRG
−1
Au NR 및 20μg mL
−1
Au@Ag NR. 아 차량 통제; ㄴ Au NR; ㄷ Au@Ag NR
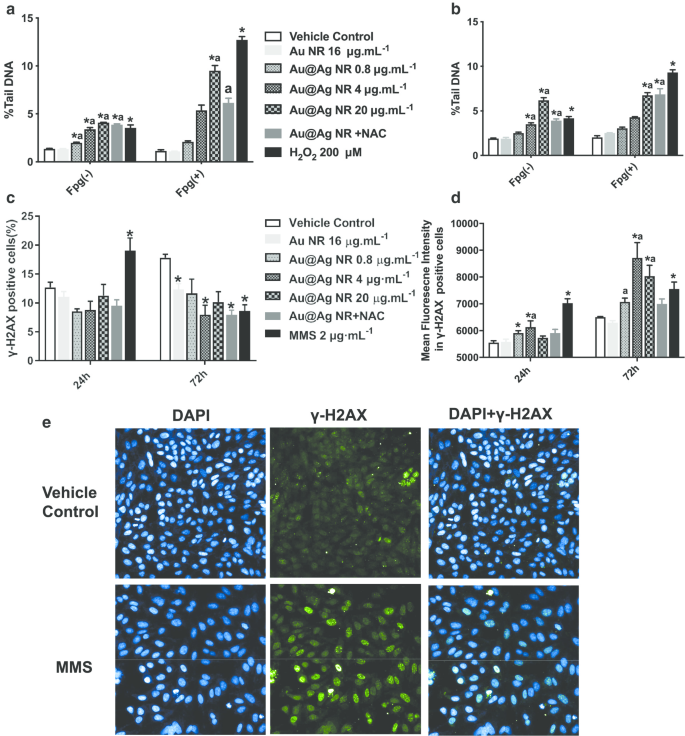
DNA 손상
Au@Ag NR에 의해 유발된 DNA 손상은 comet assay와 γH2AX assay 모두에 의해 평가되었습니다(그림 3). 혜성 분석에서 0.8 ~ 20µg mL
-1
Au@Ag NR은 상당한 DNA 손상을 일으킬 수 있습니다. Au@Ag NR에 24시간 또는 72시간 노출 후, 세포의 % 꼬리 DNA와 OTM 모두 시간 및 농도 의존적 방식으로 증가했습니다. 또한 산화 스트레스 유도와 관련된 DNA 손상이 20 µg mL
-1
로 처리된 세포에서 관찰되었습니다. Fgp 효소 변형 혜성 분석에 의한 Au@Ag NR(그림 3a, b). 암 발생과 더 높은 상관관계를 나타내는 이중 가닥 파손 정도를 평가하기 위해 γ-H2AX 양성 세포와 γ-H2AX 양성 세포의 평균 형광 강도를 모두 분석했습니다. Au@Ag NR에 24시간 노출시킨 후 γ-H2AX 양성 세포에서 그룹 간에 차이가 발견되지 않았습니다. 그러나 4μg mL
−1
Au@Ag NR 그룹은 72시간 처리 후 유의한 증가를 일으켰습니다. 비히클 대조군과 비교하여 72시간 후 모든 Au@Ag NR 그룹에서 형광 강도의 상당한 증가가 관찰되었습니다(그림 3c–e, P <0.05).
<사진>
Au@Ag NR에 의해 유도된 DNA 손상. HepaGR 세포는 다양한 농도(0.8 ~ 20μg mL
−1
)에서 Au@Ag NR에 노출되었습니다. ) 각각 24시간 및 72시간 동안 아 24시간 동안 Au@Ag NR에 노출된 후 평균 % 꼬리 DNA; ㄴ 72시간 동안 Au@Ag NR에 노출된 후 평균 % 꼬리 DNA; ㄷ 유세포 분석을 사용하여 추정된 γ-H2AX 병소를 갖는 양성 세포의 백분율; d 면역형광 염색을 사용하여 추정된 γ-H2AX 병소가 있는 세포의 평균 형광 강도.
*피 <0.05 대 차량 제어;
아피 <0.05 대 Au NR. 2μM mL
−1
MMS는 양성 대조군으로 사용되었습니다.
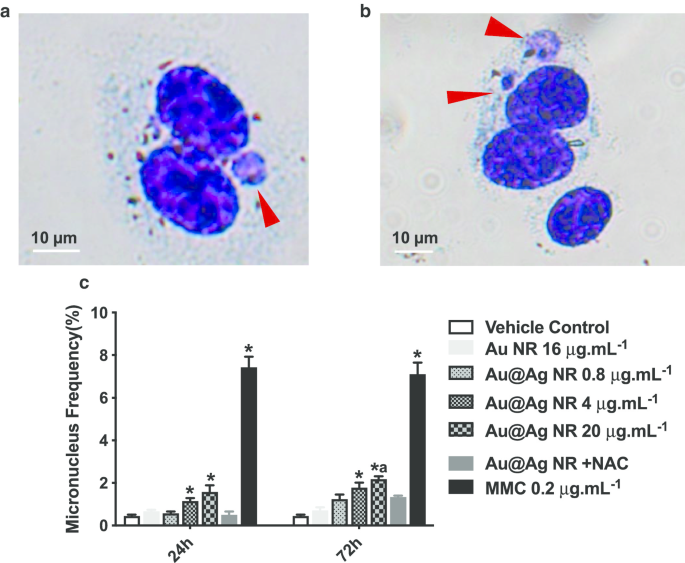
염색체 손상
소핵의 형성은 DNA 파손보다 유전 물질에 대한 더 심각한 손상인 염색체 손상을 식별하는 중요한 바이오마커입니다. 소핵을 포함하는 이핵 세포의 비율은 그림 4c와 같이 점수가 매겨졌습니다. Au@Ag NR은 농도 의존적 패턴으로 소핵 형성을 증가시켰다. 24시간 노출 후 4 μg mL로 처리한 세포에서 관찰된 소핵의 비율
-1
Au@Ag NR 및 20μg mL
−1
Au@Ag NR은 각각 1.133 ± 0.145% 및 1.567 ± 0.318%로, 둘 다 차량 대조군보다 유의하게 높았다. 72시간 노출 후 4μg mL
−1
로 처리된 세포의 소핵 비율 Au@Ag NR은 1.767 ± 0.233%로 비히클 대조군보다 유의하게 높았다. 20μg mL
−1
로 처리된 세포의 소핵 비율 Au@Ag NR은 2.167 ± 0.252%로 비히클 대조군과 16μg mL
−1
모두에서 관찰된 것보다 훨씬 높았습니다. Au NR 그룹(0.700 ± 0.153%). 대조적으로, 20μg mL
−1
로 처리된 세포 간에는 차이가 발견되지 않았습니다. Au@Ag NR + NAC 및 비히클 제어, Au@Ag NR에 의해 유도된 염색체 파손에 ROS의 참여를 시사합니다.
<사진>
Au@Ag NR에 의해 유도된 염색체 손상. HepaGR 세포는 0.8μg mL
−1
의 다양한 농도에서 Au@Ag NR에 노출되었습니다. ~ 20μg mL
−1
24시간 및 72시간 동안 아 , b 소핵의 대표적인 이미지(빨간색 화살표); ㄷ 소핵 빈도(%).
*피 <0.05 대 차량 제어;
아피 <0.05 대 Au NR. 0.2μg mL
−1
미토마이신 C를 양성 대조군으로 사용
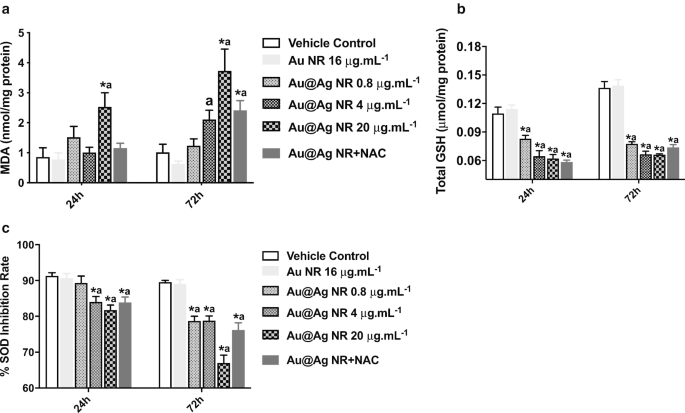
Au@Ag NR이 ROS 형성에 미치는 영향
Au@Ag NR-유도 DNA 및 염색체 손상에서 ROS 형성의 역할을 더 조사하기 위해 MDA, GSH 및 SOD 수준을 추정했습니다. MDA 형성의 상당한 증가(P <0.05)는 20μg mL에 노출된 후 관찰되었습니다.
−1
24시간과 72시간 모두에 대한 Au@Ag NR(그림 5a). 또한, Au@Ag NR에 노출된 세포의 GSH 및 SOD 수준은 상당한 감소를 보였습니다(P <0.05) 시간 및 농도 의존적 방식으로. 이러한 결과는 Au@Ag NR의 노출에 의해 생성된 산화와 항산화 사이의 불균형을 시사합니다(그림 5b, c).
<그림>
Au@Ag NR이 ROS 형성에 미치는 영향. HepaGR 세포는 0.8μg mL
−1
의 다양한 농도에서 Au@Ag NR에 노출되었습니다. ~ 20μg mL
−1
24시간 및 72시간 동안 아 MDA 수준; ㄴ GSH 수준; ㄷ SOD 수준.
*피 <0.05 대 차량 제어;
아피 <0.05 대 Au NR
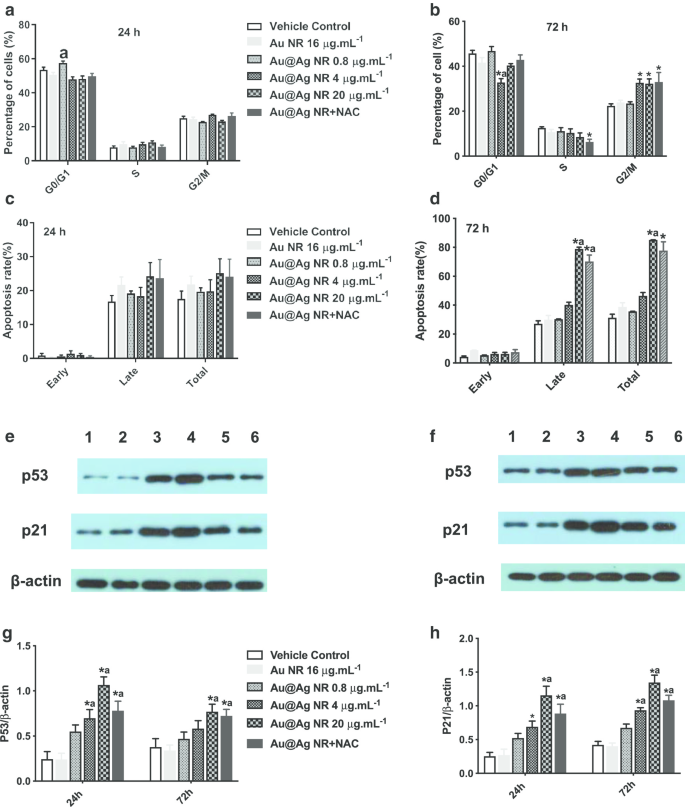
Au@Ag NR이 세포 주기 및 세포 사멸에 미치는 영향
Au@Ag NR에 72시간 노출 후 G2/M기의 세포 수 증가가 4μg mL
−1
에서 관찰되었습니다. Au@Ag NR, 20μg mL
−1
Au@Ag NR과 Au@Ag NR + NAC 그룹은 각각 32.63% ± 1.77%, 32.267% ± 2.17% 및 32.967% ± 4.25%의 비율로 b보다 각각 유의하게 컸습니다(그림). 차량 대조군(22.37% ± 0.92%). 한편, Au@Ag NR에 의해 유도된 세포 사멸은 72시간 노출 후에 관찰될 수 있었고, 20μg mL로 처리된 세포의 후기 세포 사멸 속도는
-1
Au@Ag NR 및 20μg mL
−1
Au@Ag NR + NAC는 각각 78.90 ± 1.19% 및 70.20 ± 4.50%였습니다(그림 6c, d). Au@Ag NR은 초기 세포사멸보다 후기 세포사멸을 더 많이 유도했으며, NAC의 치료는 Au@Ag NR에 의해 촉발된 후기 세포사멸의 세포율을 완화할 수 있습니다.
<그림>
Au@Ag NR이 세포 주기와 세포 사멸에 미치는 영향. Au@Ag NR이 세포 주기에 미치는 영향(a , b ) 및 세포자멸사(c , d ) 각각 24시간 및 72시간 노출 후 다른 그룹의 HepaRG 세포에서 p53 및 p21의 발현 수준에 대한 대표적인 데이터(e , f 레인 1:차량 통제; 레인 2:Au NR; 레인 3:Au@Ag NR + NAC; 레인 4:Au@Ag NR 20μg mL
−1
; 레인 5:Au@Ag NR 4 μg mL
−1
; 레인 6:Au@Ag NR 0.8μg mL
−1
); 다른 그룹에서 β-액틴에 대한 p53 및 p21의 평균 상대 발현 수준은 (g , f ).
*피 <0.05 대 차량 제어;
아피 <0.05 대 Au NR
p21과 p53의 발현 정도는 Western blot으로 확인하였고 유사한 양상을 보였다. 4μg mL
−1
로 처리한 세포에서 p53 및 p21의 발현 수준 및 20μg mL
−1
Au@Ag NR이 현저하게 증가했습니다(P <0.05) 20μg mL
−1
으로 처리된 세포에서 유의하게 감소했습니다. Au@Ag NR 및 NAC(P <0.05, 20μg mL와 비교
−1
Au@Ag NR 그룹, 그림 6e–h). p53 단백질은 DNA 손상에 대한 반응으로 G2/M 체크포인트 활성화를 매개하는 핵심 분자로 알려져 있으며, p21은 p53 의존적 세포주기 억제제로 인식되고 있다. 따라서 Au@Ag NR은 DNA 복제를 방해하고 세포 주기 정지에 의해 DNA 복구를 방해할 수 있습니다.
토론
현재 출시된 Ag
+
의 역할은 유전 독성을 생성하는 AgNPs는 분명하지 않습니다. 우리 그룹[21]과 다른 그룹[13]의 이전 연구는 Ag
+
독성을 도입하는 주요 원인이며, 나노입자도 매우 독성이 있을 수 있습니다. 예를 들어, AgNPs는 하이드록실 라디칼의 형성을 유도하여 유전독성에 기여할 수 있습니다[13]. 또한 Ag
+
에 비해 AgNP에 의해 더 심각한 염색체 손상, 산화 스트레스 및 세포 사멸이 도입되었습니다. 단독으로 [23], 다른 경로가 관련될 수 있음을 시사합니다. Au@Ag NR을 세포 내 AgNPs의 형태와 분포를 이해하기 위한 모델 재료로 사용했으며, ICP-MS로 세포 내 Ag 및 Au의 양을 결정했습니다. 준비된 Au@Ag NR의 Ag/Au 중량비는 2.3으로 추정되었다. However, after a 24-h exposure, it sharply increased to 16.5 in the cells treated with Au@Ag NR, suggesting that large amount of Ag was released from the shell of Au@Ag NR within that period. When the exposure period of Au@Ag NR was extended to 72 h, the Au/Ag weight ratio was decreased to 1.7, indicating that the Ag
+
was released from the cell and the nanorod was the major form of Au@Ag NR in the cell at that stage. Therefore, it could be deduced that once the Au@Ag NR entered the cell, Ag
+
rapidly dissolved from its shell within 24 h and gradually released to the extracellular environment, while the Au@Ag NR itself retained in the cell for a longer period.
Oxidative stress is deemed as one of the most important toxicological mechanisms of nanoparticles [24]. N-acetylcysteine (NAC) is a thiol, a mucolytic agent and precursor of l-cysteine which reduced glutathione. NAC is also a source of sulfhydryl groups in cells and exerts the ROS scavenger activity by interacting with OH·
및 H2 O2 [25]. In this study, the GSH and SOD levels were significantly decreased after exposure to Au@Ag NR, while the MDA level increased in a concentration- and time-dependent manner, indicating that the Au@Ag NR introduced the oxidative stress in the cells.
The potentials of Ag and Au@Ag NR in interfering with the genetic materials were further investigated by a series of genotoxicity assays. It is noteworthy that co-culturing the NAC with Au@Ag NR could ameliorate the ROS formation, which in turn supports the participation of oxidative stress in the genotoxicity triggered by Au@Ag NR. In this study, comet and γ-H2AX assays were performed to confirm that Au@Ag NR could interact with DNA and induce certain DNA damage, and the repair endonuclease Fpg was included in the comet assay to identify the oxidative DNA damage [26]. The Fgp could recognize oxidized pyrimidines and remove oxidized purines, e.g., 8-hydroguanine, so as to create apurinic or apyrimidinic sites that could introduce gaps in the DNA strands. The oxidative stress-induced DNA breakage could be determined subsequently by another comet assay [27]. The further DNA breakage detected by the additional Fgp in the comet assay suggested that the Au@Ag NR could cause DNA damage. Mei et al. [28] observed that 5-nm-sized AgNPs induced oxidative lesion-specific DNA damage by employing the hOGG1, EndoIII and Fpg endonucleases in the comet assay. Li et al. [29] also suggested that both PVP- and silica-coated AgNPs (15–100 nm and 10–80 nm, respectively) could lead to a significant increase in DNA breakage in mice hepatocytes in the presence of hOGG1and EndoIII. The formation of γ-H2AX foci, which represents an early cellular response to genotoxic stress, is the most sensitive and specific biomarker for detecting DSBs [30]. As demonstrated in this study, γ-H2AX foci in cells exposed to Au@Ag NR were markedly increased after 24 h, and a further increase could be observed after 72 h. The reduction in the 20 µg mL
−1
group might be due to the cytotoxicity to the HepaRG cells at higher concentration. Similar results were observed for AgNPs with different coatings [31, 32]. Further, our results suggest that Au@Ag NR could induce chromosome damage in HepaRG cells, as the micronucleus rates were significantly increased. This is consistent with previous studies, where AgNPs-induced increased micronucleus rate was reported in HaCaT and TK6 cells [33]. In contrast, the addition of oxidative radical scavenger NAC could inhibit the formation of micronucleus induced by Au@Ag NR. Taken together, these data suggest the participation of oxidative stress in AgNP-introduced clastogenicity risk in vitro.
Previous studies have investigated the cell cycle arrest and cytotoxicity induced by AgNPs [33,34,35]. With prolonging the exposure time, the impact of AgNPs on cell cycle and apoptosis might be enhanced and in turn aggravate the cytotoxicity and genotoxicity. Usually, the cell cycle checkpoints (e.g., G2/M) were initiated by cells when experiencing DNA damage, and this mechanism serves to prevent the cell from entering mitosis (M phase). The G2/M cell cycle arrest indicates that an increasing percentage of cells is hindered in G2 phase for DNA repairing. Cells experiencing successful DNA repairing would further proceed to mitosis; however, for those with fatal damages, irreversible G2/M cell cycle arrest and cells apoptosis would take place [36]. We observed that Au@Ag NR could arrest the majority of HepaRG cells in G2/M phase, induce late cell apoptosis and increase the expression levels of p53 and p21, which are important proteins associated with the regulation of cell cycles [37]. As p53 could also induce apoptosis, when the DNA cannot be repaired properly [38], the p21 might indirectly participate in cell apoptosis by cell cycle arrest in a p53-dependent pathway via down-regulating the nuclear protein ICBP90 for DNA replication and cell cycle regulation [39]. Furthermore, apoptosis and a G2/M arrest induced by activation of the p53/p21 system have been reported in HepG2 cells following the administration of garlic extracts [40]. Thus, it could be inferred that the oxidative stress-triggered DNA/chromosome damages might facilitate the expression of p53 and p21, which subsequently induces cell cycle arrest. Extending the exposure period of Au@Ag NRs to the DNA/chromosome during replication may further aggravate the genotoxicity or apoptosis.
Conclusion
Genotoxicity induced by AgNPs may be attributed to the oxidative stress induced by the nanoparticles as well as the released ions [41]. This study employed Au@Ag NR as a model to determine the distribution and release behavior of Ag after the nanoparticles enter into the cells. Considering the disparate forms of Au@Ag NR in the cell, after its exposure the Ag
+
was rapidly dissolved from the silver shell. Ag
+
and Au@Ag NR could introduce cytotoxicity and genotoxicity (clastogenicity) in the cells, and the Au@Ag NR retained in the nucleus may further release Ag
+
to aggravate the damage, which are mainly caused by cell cycle arrest and ROS formation (summarized in Fig. 7). Collectively, these data reveal the correlation between the intracellular accumulation, Ag
+
release as well as the potential genotoxicity of AgNPs.
Schematic diagram of the possible mechanism of genotoxicity introduced by AgNP in vitro