나노 입자 과학은 다양한 과학 분야의 지형을 빠르게 변화시키고 새로운 기술 플랫폼을 정의하고 있습니다. 이것은 아마도 나노입자가 많은 질병의 치료 및 진단을 위한 도구로 사용되어 온 나노의학 분야에서 더욱 분명해질 것입니다. 그러나 엄청난 이점이 있음에도 불구하고 이 기술의 일반적인 함정은 인체에 대한 잠재적인 장단기 영향입니다. 이러한 문제를 이해하기 위해 많은 과학적 연구가 수행되었습니다. 이 검토는 이러한 연구와 그 결과 중 일부를 조명하려고 시도합니다. 이 리뷰에서 조사한 주제에는 나노 입자의 다양한 가능한 흡수 경로와 세포 내 인신 매매 경로가 포함됩니다. 또한, 나노입자의 흡수 메커니즘 및 생물학적 기능을 결정하는 데 있어 크기, 모양, 전하 및 표면 화학과 같은 나노입자의 물리화학적 특성의 영향도 설명합니다.
소개
나노입자(NP)는 고유한 특성과 엄청난 적용 가능성으로 인해 상상할 수 있는 거의 모든 분야에서 현재 최첨단 연구의 최전선에 있는 나노 물질의 하위 범주입니다[1,2,3,4]. RNCOS의 "Global NP Market Outlook 2020"이라는 기술 시장 조사 보고서에서 NP 시장은 2015-2020년 동안 16%의 CAGR(연간 복합 성장률)로 성장할 것으로 보고되었습니다. NP 기술은 빠르게 성장하고 있는 응용 레퍼토리와 함께 생물 의학 및 생명 공학 분야에서 독특한 틈새 시장을 찾았습니다[5, 6]. 예를 들어, NP는 약물 및 유전자 전달[7, 8] 병원체의 생체 탐지[9], 단백질 탐지[10], 조직 공학[11, 12], 종양 영상화 및 표적화[13], 고열[14] 및 MRI 조영증강[15].
크기가 작기 때문에 NP는 세포에 쉽게 들어갈 수 있을 뿐만 아니라 세포, 조직 및 기관을 가로질러 이동할 수 있습니다. NP는 생물학적 장벽을 통과하고 세포에 들어가 기능을 발휘할 수 있기 때문에 생물 의학 응용 분야에서 널리 사용됩니다. 그러나 양날의 검처럼 NP의 잠재적 위험(즉, 역효과)도 이 능력에서 발생합니다[16, 17]. "작은" 크기에도 불구하고 극성 분자인 NP는 세포막(CM)을 통해 확산될 수 없습니다. CM은 대부분 작고 비극성인 분자에 투과성이기 때문에 NP는 세포 내로 들어가기 위해 세포내이입 경로를 사용합니다[18, 19]. 나노입자가 세포에 들어가는 방식은 생체의학적 기능, 생체분포 및 독성을 결정하는 핵심 요소입니다. 나노의학에서 나노입자의 세포 내로의 안전한 진입은 높은 치료 효능을 얻기 위한 중요한 단계입니다. 또한, NP의 세포 내 트래피킹 및 운명은 이러한 운반체가 특정 세포하 구획을 표적으로 하고 조영제, 유전자 및 약물과 같은 특정 생체 분자를 전달하는 것을 목표로 한다는 점을 고려하면 NP의 성공에 중요한 과정입니다[18, 20,21,22 ]. 더 중요하게는, NP에 의한 세포독성의 유도는 진입 경로 및 세포내 국소화에 의해 결정됩니다. 따라서 NP의 세포 흡수 및 세포 내 트래피킹을 이해하는 것은 안전하고 효율적인 나노 의약품을 설계하는 데 중요합니다[23].
NP의 세포 흡수, 표적화 및 세포 내 트래피킹은 크기, 모양 및 표면 특성과 같은 NP의 물리화학적 특성을 조정하여 최적화할 수 있습니다[24]. 따라서 세포 흡수와 관련된 기본 메커니즘에 대한 지식은 NP의 운명과 독성을 평가하는 데 중요합니다. 이 검토는 NP의 다양한 가능한 흡수 경로와 세포 내 인신 매매 경로를 강조합니다. 또한 크기, 모양, 전하 및 표면 화학과 같은 NP의 물리화학적 특성이 세포 내재화에 미치는 영향도 설명합니다. 세포 흡수 메커니즘과 관련하여 NP의 물리화학적 특성을 이해하면 주변의 건강한 조직에 대한 독성 영향을 최소화하면서 제어된 방식으로 표적 작용 부위에 약물 페이로드를 전달하는 것과 같은 생의학 응용 분야에서 중요한 기능적 NP를 설계할 수 있습니다. 및 장기.
NP의 세포 흡수 경로
원형질막이라고도 하는 CM은 세포외액에서 세포내를 분리하여 세포질을 둘러쌉니다. CM은 세포 내 구성 요소를 보호하고, 세포 항상성을 유지하고, 구조적 지원을 제공하고, 세포 구성을 유지하기 때문에 매우 중요합니다[25,26,27,28,29]. CM은 단백질이 포함된 이중층으로 배열된 인지질로 구성됩니다. 친수성 머리와 소수성 꼬리를 가진 이러한 인지질 이중층은 작은 생체 분자의 진입을 허용합니다. 보다 구체적으로, CM은 세포로 물질의 통과를 제어하는 선택적 투과성 장벽입니다[30, 31]. CM은 주로 수동 수송과 능동 수송의 두 가지 범주로 구분되는 물질을 교환하기 위해 다른 메커니즘을 사용합니다. 산소 및 이산화탄소와 같은 가스, 벤젠과 같은 소수성 분자, 물 및 에탄올과 같은 전하를 띠지 않는 분자는 농도가 높은 영역에서 낮은 영역으로 막을 가로질러 확산됩니다. 농도 구배를 따라 에너지의 도움 없이 발생하는 이러한 종류의 수송을 수동 수송이라고 합니다. 대조적으로, 활성 수송은 ATP(adenosine triphosphate)가 제공하는 에너지를 사용하여 농도 구배에 대해 발생합니다[32,33,34,35,36].
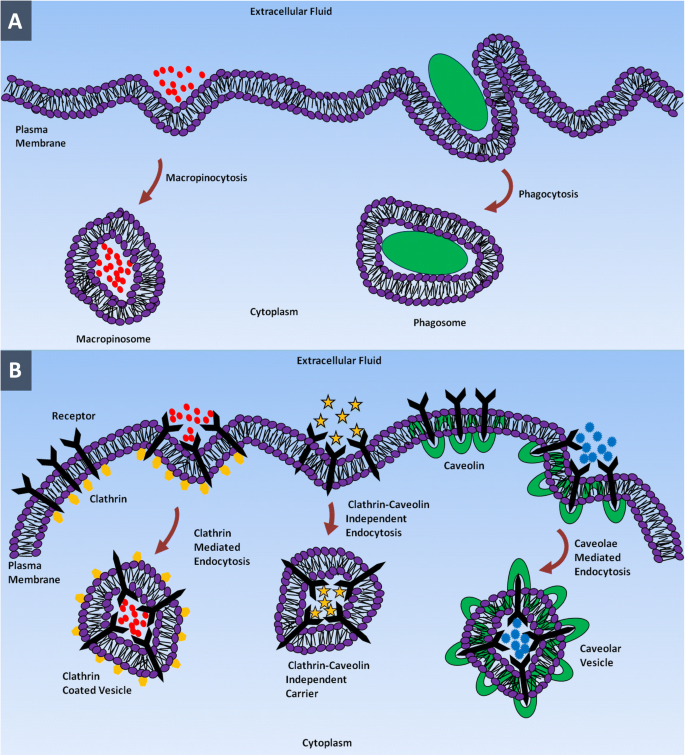
소수성 원형질막을 통과할 수 없는 극성 또는 하전된 생체 분자는 세포내이입(endocytosis)이라고 하는 능동 수송 형태에 의해 내부화됩니다. 이 과정에서 세포는 CM의 함입에 의해 세포외액 내부의 물질을 삼키고 세포 내부에서 싹이 트면서 엔도솜이라고 하는 막으로 둘러싸인 소포를 형성합니다[37]. Endocytosis는 기본적으로 phagocytosis와 pinocytosis의 두 가지 주요 범주로 분류할 수 있습니다. 식세포 작용(세포 섭취)은 식세포(즉, 단핵구, 대식세포 및 호중구)라고 하는 특수 포유동물 세포에 의해 파편, 박테리아 또는 기타 큰 크기의 용질을 섭취하는 과정입니다[38, 39].
식세포 작용에 필수적인 것은 면역 글로불린 및 보체 단백질과 같은 옵소닌이 표적 물질을 코팅하여 식세포의 존재를 촉발하고 식세포 활성을 초기화하는 옵소닌화라고 하는 과정입니다[40]. 식세포가 표적 물질을 섭취하기 시작하면 동시에 식세포 내에서 섭취된 물질이 구획화되는 식세포(phagosome)라고 하는 막 결합 소포의 형성을 자극합니다. 이 과정의 마지막 단계에서 phagosome은 리소좀과 융합하고 물질은 리소좀 내강에 포함된 가수분해 효소에 의해 산성 pH에서 소화됩니다[41,42,43].
모든 세포 유형에서 나노미터 범위의 작은 입자는 음세포 작용에 의해 내부화됩니다[44]. 피노사이토시스에서 "세포 음주" 원형질막은 그 안에 용해된 분자를 포함하는 세포외액의 작은 방울을 흡수하기 위해 함입을 형성합니다. Pinocytosis는 구별하는 과정이 아니며 세포의 필요에 관계없이 거의 모든 세포에서 연속적으로 발생합니다. 포획된 물질은 리소좀과 융합하여 내용물을 가수분해하거나 분해하는 피노좀이라고 하는 작은 소포로 꼬집어집니다[45, 46]. 식균 작용과 음세포 작용은 세포 내이입 소포의 크기로 구분할 수 있습니다. 전자는 250 nm 크기의 큰 소포에 의한 큰 입자의 흡수를 포함하고, 후자는 크기가 수 나노미터에서 수백 나노미터 범위인 작은 소포를 통한 유체의 흡수를 포함합니다[42, 47]. Pinocytosis는 clathrin-매개 endocytosis, caveolae-매개 endocytosis, clathrin 및 caveolae-independent endocytosis 및 macropinocytosis로 세분될 수 있습니다[48, 49].
클라트린 매개 엔도사이토시스는 특정 분자를 세포 내로 내재화하는 세포 진입 메커니즘입니다. 이 진입 경로는 세포가 저밀도 지단백질 수용체에 의해 콜레스테롤을 포함한 원형질막 성분 및 영양소를, 트랜스페린 수용체에 의해 흡수되도록 돕습니다[50,51,52,53,54,55,56]. 이 과정에서 세포외액의 특정 리간드는 CM 표면의 수용체에 결합하여 리간드-수용체 복합체를 형성합니다. 이 리간드-수용체 복합체는 clathrin이 풍부한 CM의 특수 영역으로 이동하여 clathrin-coated vesicle의 형성을 통해 삼켜집니다. 일단 세포 내부에 들어가면 소포 외부의 클라트린 코팅이 초기 엔도솜과 융합하기 전에 배출됩니다. 초기 엔도좀 내의 화물은 결국 엔도-리소좀 경로를 통해 리소좀에 도달합니다[40, 57,58,59,60]. 각 유형의 NP는 우선적으로 흡수 경로를 통해 세포에 의해 내재화됩니다. 예를 들어, 폴리(락트산-코-글리콜산), D,L-폴리락타이드 및 폴리(에틸렌 글리콜코-락타이드) 및 실리카로 구성된 NP(SiO2 ) 기반 나노물질은 클라트린 매개 세포내이입 경로에 의해 내재화된다[61]. 쿠마린 기반 고체 지질 NP는 이러한 NP의 구조가 CM과 유사하기 때문에 비에너지 의존성 경로를 통해 세포에 의해 내재화됩니다. 모든 지질 기반 나노입자는 클라트린 매개 엔도사이토시스 경로를 이용합니다[62]. 허셉틴으로 코팅된 금 나노입자는 막 ErbB2 수용체를 통해 수용체 매개 세포내이입을 통해 세포로 들어갑니다[63].
Caveolae 매개 endocytosis는 caveolae(작은 동굴)라고 하는 플라스크 모양의 막 함입을 포함하는 세포 진입 경로입니다. Caveolae는 내피 세포, 상피, 지방 세포, 근육 및 섬유 아세포 세포에 존재합니다 [64,65,66,67]. 동굴의 크기는 일반적으로 50~80 nm 범위이며 플라스크 모양의 구조를 부여하는 막 단백질 카베올린-1로 구성됩니다[68,69,70,71]. Caveolae-dependent endocytosis는 세포 신호 전달 및 막 단백질, 지질 및 지방산의 조절에 관여합니다[61, 64, 67]. 동굴형이 원형질막에서 분리되면 중성 pH에 존재하는 동굴형이라는 세포 구획과 융합됩니다. Caveosomes는 리소좀을 우회할 수 있으므로 가수분해 효소와 리소좀 분해로부터 내용물을 보호합니다. 따라서 바이러스 및 박테리아를 포함한 병원체는 이 진입 경로를 사용하여 분해를 방지합니다. 카베올린 의존성 기전에 의해 세포로 내재화된 화물은 리소좀으로 끝나지 않기 때문에 이 경로는 나노의학에서 사용된다[54, 72,73,74].
클라트린 및 카베올린 독립적 엔도사이토시스는 클라트린과 카베올린이 결핍된 세포에서 발생합니다. 이 경로는 성장 호르몬, 세포외액, 글리코실포스파티딜이노시톨(GPI) 연결 단백질 및 인터루킨-2가 세포로 들어가는 데 활용됩니다. 예를 들어, clathrin 및 caveolae 독립적인 경로를 사용하여 세포에 들어가는 엽산[58, 72, 75,76,77,78,79]은 약물 전달 시스템 및 영상화제로 사용되는 NP 및 폴리머에 접합됩니다[53 , 80, 81]. Macropinocytosis는 세포가 macropinosomes라는 큰 소포(0.5-10μm)를 형성하여 많은 양의 세포외액을 흡수하는 일종의 pinocytosis 메커니즘입니다[82,83,84,85]. Macropinocytosis는 세포 사멸 및 괴사 세포, 박테리아 및 바이러스뿐만 아니라 항원 제시를 내부화하는 경로입니다. 이 경로는 대부분의 다른 경로로는 세포로 들어갈 수 없는 마이크론 크기의 NP를 내재화할 수 있습니다. 거대음세포증은 뇌 미세혈관 내피세포를 제외한 거의 모든 세포에서 발생할 수 있다[86,87,88,89]. NP는 그림 1과 같이 이러한 세포내이입 경로 중 하나를 통해 세포로 들어갑니다.
<그림>
다른 세포 내 이입 경로를 사용하여 NP가 세포로 진입합니다. 아 Macropinocytosis 및 phagocytosis. ㄴ Clathrin 매개 endocytosis, clathrin-caveolin 독립 endocytosis 및 caveolae 매개 endocytosis
세포 흡수에 대한 NP의 물리화학적 특성 효과
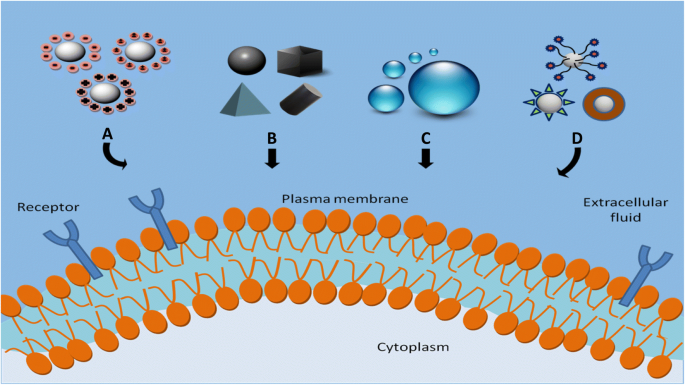
크기, 모양, 표면 전하, 표면 소수성/친수성 및 세포 흡수에 대한 표면 기능화와 같은 NP의 물리화학적 특성의 효과를 연구하는 것은 이러한 매개변수가 NP의 흡수 수준, 세포내이입 경로 및 세포독성에 직접적인 영향을 미치기 때문에 중요합니다. [90, 91]. NP의 세포 흡수에 영향을 미치는 물리화학적 요인은 그림 2에 나와 있습니다. 다음 섹션에서는 이러한 매개변수가 세포-NP 상호작용에 미치는 영향에 대해 설명합니다.
<그림>
NP의 세포 흡수에 영향을 미치는 물리화학적 요인. 아 표면 전하, b 모양, c 크기 및 d 표면 화학
크기의 영향
NP의 크기는 세포 흡수[92]의 효율성과 살아있는 세포에 대한 독성 잠재력[24]을 결정하는 핵심 요소입니다. 또한, NP의 크기도 흡수 경로를 결정하는 데 중요한 역할을 하는 것으로 밝혀졌습니다. 크기가 수에서 수백 나노미터에 이르는 작은 NP는 음소 또는 대음포작용을 통해 세포에 들어갑니다. 250 nm에서 3 μm 크기 범위의 NP는 최적의 시험관 내 식균 작용을 갖는 것으로 나타났지만, 120-150 nm 크기 범위의 NP는 클라트린 또는 카베올린 매개 엔도사이토시스를 통해 내재화되고 NP의 최대 크기 이 경로를 사용하는 것은 200 nm인 것으로 보고되었습니다[47, 93]. 동굴 매개 경로에서 동굴의 크기는 더 큰 나노입자의 흡수를 방해합니다[16, 17]. 특정 유형의 NP는 크기에 따라 여러 세포내이입 경로를 사용할 수 있습니다.
여러 연구에서 NP의 세포 흡수를 위해 NP가 더 효율적으로 내재화되고 더 높은 흡수율을 갖는 50nm의 최적 크기가 있음을 나타냅니다. NP 흡수는 더 작은 입자(약 15-30 nm) 또는 더 큰 입자(약 70-240 nm)에서 감소하는 것으로 나타났습니다[94,95,96,97,98,99]. 또한, 30-50 nm 크기의 나노입자는 CM 수용체와 효율적으로 상호작용하고 이후에 수용체 매개 세포내이입을 통해 내재화됩니다[97]. 나노입자의 약물 전달 응용에서 주요 관심사는 나노입자가 세망내피계에 의해 제거되는 것을 방지하고 혈액 내 순환 시간을 연장하여 표적에서의 생체이용률을 높이는 것입니다. 이와 관련하여 NP의 크기를 늘리면 제거율이 증가하게 됩니다[100,101,102,103,104,105]. 따라서 세포 흡수에서 NP 크기의 역할을 이해하는 것은 의료 적용을 위한 효과적이고 안전한 NP를 설계하는 데 중요합니다.
여러 연구에서 NP의 크기와 섭취 경로 사이의 관계를 조사했지만 밝혀진 결과는 항상 일관성이 없었습니다[93, 106,107,108,109]. 이러한 모순은 크기를 제어하는 과정에서 NP의 다른 매개변수를 제어하는 복잡성과 관련될 수 있습니다. 또한 합성 후 측정된 나노입자의 크기는 시험관 내 및 생체 내 연구 동안 응집 및 응집으로 인해 변화를 겪을 수 있으며, 이는 차례로 세포 내재화 경로에 영향을 미칠 수 있습니다[110, 111]. non-phagocytic B16 세포에서 입자 크기가 세포 흡수 경로에 미치는 영향은 50-1000 nm 범위에서 다양한 크기의 형광 라텍스 비드를 사용하여 조사되었습니다[93]. 결과는 이러한 비드의 내재화 메커니즘이 입자 크기에 크게 의존한다는 것을 입증했습니다. 특히, 200 nm 이하의 크기를 갖는 비드는 clathrin으로 코팅된 구덩이에 의해 흡수된 반면, 더 큰 비드는 동굴-매개 엔도사이토시스에 의해 내재화되었다. Lai와 동료[16]는 크기가 25nm 미만인 작은 고분자 나노입자가 엔도/리소좀 경로 외부의 비분해성 소포를 통해 세포의 핵주위 영역에 도달하는 새로운 메커니즘을 사용한다는 것을 발견했습니다. 이 경로는 non-clathrin 및 non-caveolae-매개 및 콜레스테롤 독립적입니다.
SK-BR-3 세포에 의해 Herceptin-AuNP와 접합된 다양한 크기(2 ~ 100 nm)의 금(Au) NP의 흡수는 크기 의존적인 것으로 나타났습니다. 가장 높은 세포 내재화는 25-50 nm 크기 범위의 NP에서 관찰되었습니다 [63]. 이 진입 경로에서 NP의 크기는 막 수용체의 결합 및 활성화와 단백질의 최종 발현에 결정적인 영향을 미치는 것으로 밝혀졌습니다. 세포내 흡수에 대한 콜로이드성 AuNP의 크기와 모양의 변화 효과가 평가되었다[112]. 구형 및 막대 모양의 14-, 50- 및 74nm 크기의 AuNP를 HeLa 세포와 함께 배양했습니다. NP 흡수율은 입자의 크기와 모양에 따라 크게 달라지며 50nm 크기의 입자가 가장 높은 흡수율을 나타냄을 알 수 있었다. 더욱이, 구형 AuNP의 흡수는 유사한 크기의 막대형 NP보다 500% 더 많았다. Shanet al. [113]은 HeLa 세포에 의해 직경이 4, 12 및 17 nm인 AuNP를 세포내이입하는 크기 의존적 힘을 조사했습니다. 결과는 AuNP의 크기에 따라 흡수 및 결합 해제 힘 값이 모두 증가하는 것으로 나타났습니다. SiO2의 흡수 A549 세포(폐 상피 세포)에 의한 다양한 크기(50, 100 및 300 nm)의 NP는 유세포 분석, 형광 및 전자 현미경의 조합을 통해 연구되었습니다. 이 연구자들은 SiO2의 흡수가 NP는 [114] 크기만큼 감소했습니다.
모양 효과
크기 외에도 NP의 모양은 NP의 수송뿐만 아니라 흡수 경로에서 중추적인 역할을 합니다. Chithraniet al. [112]는 HeLa 세포의 흡수에 대한 콜로이드 AuNP의 모양 효과를 연구했습니다. 결과는 구형 AuNP가 막대 모양의 AuNP보다 5배 더 높은 흡수율을 갖는 것으로 나타났습니다. 다른 연구에서 동일한 연구자들은 세 가지 다른 세포주에서 구형 및 막대 모양의 트랜스페린으로 코팅된 AuNP의 흡수 수준을 조사했습니다. STO 세포, HeLa 세포 및 SNB19 세포 [94]. 그들은 구형 AuNP가 막대 모양의 AuNP보다 빠른 속도로 모든 세포주에 의해 내재화된다는 것을 관찰했습니다.
생체 내에서 모양의 효과를 확인하기 위해 Geng와 동료[115]는 필로미셀을 사용하여 설치류에서 구체가 있는 유연한 필라멘트의 수송 및 수송의 차이를 평가했습니다. 결과는 필로미셀이 구형보다 약 10배 더 순환에 남아 있음을 보여주었습니다. 더욱이, 구형 필로미셀은 더 긴 필라멘트보다 더 쉽게 세포에 의해 내부화된다. Gratton과 동료[106]는 단분산 하이드로겔 입자의 모양이 HeLa 세포로의 흡수에 미치는 영향을 입증했습니다. 그들은 막대 모양의 NP가 구, 실린더 및 큐브에 비해 가장 높은 내재화 속도를 가짐을 발견했습니다. 또 다른 연구에서는 Caco-2 세포에 디스크 모양, 구형 및 막대 모양의 폴리스티렌(PS) NP를 사용하여 NP 모양이 세포 흡수에 미치는 영향을 조사했습니다. 결과는 막대와 디스크 모양의 NP가 구형 NP보다 2배 더 높게 내재화되었음을 보여주었다. 그들은 나노입자의 형태를 고려함으로써 나노입자 매개 약물 전달이 진전될 수 있다고 결론지었다[116].
Xu와 동료[117]는 육각형 시트(가로 너비 50~150nm 및 너비 10~20nm)와 같은 다양한 형태의 FITC(fluorescein isothiocyanate)를 사용하여 층상 이중 수산화물(LDH) 나노입자를 준비하여 모양이 세포 흡수에 미치는 영향을 연구했습니다. 두께) 및 막대(폭 30–60 nm 및 길이 100–200 nm). 모든 형태는 clathrin 매개 endocytosis를 통해 채택되었습니다. LDH-FITC 나노구는 세포질에 유지된 반면 LDH-FITC 나노막대는 미세소관에 의해 핵 쪽으로 이동되었다. Dasguptaet al. 세포 흡수에 대한 NP의 모양의 역할을 조사하기 위해 시뮬레이션을 적용했습니다. 그들은 nanorod 및 nanocube 모양의 NP의 멤브레인 랩핑을 시뮬레이션했습니다. 막대 모양 입자의 경우 작고 높은 포장 분율을 가진 안정적인 세포내이입 상태를 발견했습니다. 종횡비의 증가는 완전한 래핑에 바람직하지 않습니다. Nangia와 Sureshkumar [119]는 고급 분자 역학 시뮬레이션 기술을 적용하여 NP의 전위 속도에 대한 모양의 영향을 컴퓨터화했습니다. 연구의 주요 계시는 원뿔형, 큐브형, 막대형, 쌀형, 피라미드형 및 구형 NP의 전위율의 상당한 변화입니다.
표면 전하의 영향
NP의 세포 흡수에 영향을 미치는 또 다른 중요한 요소는 표면 전하입니다. 최근 10년 동안 나노 표면 개질은 NP의 표면 전하가 양이온 또는 음이온이 되도록 조작하기 위해 사용되었습니다[92]. 음으로 하전된 CM은 양으로 하전된 NP의 흡수를 향상시킵니다. 특히, 양으로 하전된 NP는 중성 및 음으로 하전된 NP보다 더 높은 내재화를 갖는다[47, 120]. 그러나 양전하를 띤 NP의 흡수는 CM의 완전성을 방해하고 독성을 증가시킬 수 있습니다[121, 122]. 일반적으로 양전하를 띤 나노입자는 세포사를 유도한다[123, 124]. 흥미롭게도, 중성으로 하전된 NP는 음으로 하전된 NP에 비해 세포 흡수를 낮춥니다[110, 125,126,127]. 더욱이, 음으로 하전된 NP의 내부화는 막의 겔화로 이어지는 반면, 양으로 하전된 NP는 CM에서 유동성을 유발합니다[128, 129]. NP의 흡수율 외에도 표면 전하는 흡수 메커니즘에 영향을 미칩니다. 보다 구체적으로, 양전하를 띤 NP는 주로 거대음세포작용을 통해 세포에 의해 내재화되는 반면, 클라트린-/카베올라-독립 엔도사이토시스는 음전하를 띤 NP의 흡수 메커니즘이다[130]. AuNPs의 표면이 유기 분자에 의해 코팅될 때 세포 흡수 경로는 다양합니다. 예를 들어, 양으로 하전된 일반 AuNP는 거대음세포작용과 클라트린 및 카베올린 매개 엔도사이토시스를 통해 내재화되는 반면, 음으로 하전된 폴리에틸렌 글리콜(PEG) 코팅된 AuNP는 주로 카베올린 및/또는 클라트린 매개 엔도사이토시스를 통해 내재화됩니다[131]. /P>
Li와 Gu [132]는 분자 역학 시뮬레이션을 통해 하전 및 중성 NP와 CM의 상호 작용을 연구했습니다. 하전 된 NP는 중성 NP에 비해 CM에 더 나은 접착력을 갖는 것으로 밝혀졌습니다. 또한 NP의 전하 밀도를 증가시켜 NP를 멤브레인으로 완전히 감쌀 수 있습니다. 다른 연구 그룹은 양이온 및 음이온 AuNP와 CM의 상호 작용을 조사하기 위해 분자 역학 시뮬레이션을 사용했습니다. 결과는 AuNPs의 전하 밀도가 향상됨에 따라 AuNPs 침투로 인한 CM에 대한 파괴가 증가함을 보여주었다[133]. 이러한 발견은 AuNP의 표면 전하 밀도를 조작하여 AuNP의 흡수를 최적화하는 동시에 세포와 AuNP 사이의 상호 작용을 제어하는 방법을 제안하는 동시에 생물 의학 응용 분야에서 고려되는 모든 NP의 필수 특성인 세포 독성을 최소화합니다.
Li와 Malmstadt[134]는 양전하 및 음전하를 띤 PS-NP와 생물학적 막의 상호작용을 연구했습니다. 결과는 양이온성 NP와 막의 인산염 그룹 사이의 강한 정전기적 상호작용이 NP-막 결합과 막 표면 장력을 향상시켜 기공의 형성을 초래한다는 것을 보여주었다. SK-BR-3 세포로의 양전하 AuNP의 흡수율은 음전하 AuNP보다 5배 더 높은 것으로 보고되었습니다. 이 연구자들은 또한 양전하를 띤 AuNP가 비내포작용 경로에 의해 내재화되는 반면 음전하를 띤 AuNP는 세포내이입 경로를 통해 세포에 흡수된다는 사실을 탐구했습니다[135].
Hauck et al. [107]은 HeLa 세포에 의해 크기 범위가 18~40nm이고 표면 전하가 + 37mV~- 69mV인 금 나노막대(AuNR)의 흡수를 조사했습니다. 결과는 AuNR의 모든 농도에서 HeLa 세포로의 가장 높은 내재화가 + 37 mV의 표면 전하와 - 69 mV에서 가장 낮은 내재화임을 나타냅니다. Huhn과 동료[136]는 3T3 섬유아세포, 쥐 C17.2 신경 전구 세포 및 인간 제대 정맥 내피 세포와 같은 다른 세포주와 콜로이드성 AuNP의 전하 의존적 상호작용을 평가했습니다. 결과는 모든 세포주에 대해 양이온성 AuNP가 음이온성 대응물보다 더 높은 흡수를 갖는다는 것을 보여주었다. 그들은 세포 흡수가 전하의 부호에 크게 의존한다고 결론지었습니다. 또한, 세포 독성 연구는 양전하를 띤 NP에 대한 더 높은 흡수의 결과로 음전하를 띤 NP보다 더 높은 독성을 나타냅니다.
소수성의 효과
NP의 소수성은 CM과의 상호작용에서 결정적인 요소입니다[92, 137]. 여러 연구는 CM과의 상호 작용에 대한 NP의 소수성의 영향을 보여주었습니다. Li et al. [138]은 분자 역학 시뮬레이션을 사용하여 CM과의 상호 작용에 대한 NP의 소수성/친수성 효과를 연구했습니다. 결과는 소수성 NP가 CM에 포함을 생성하는 반면 친수성 NP는 CM에 흡착하는 것으로 밝혀졌습니다. 다른 연구에서는 NP-세포 상호작용에 대한 소수성의 영향을 조사하기 위해 시뮬레이션 접근이 적용되었습니다. 친수성 나노입자는 감싸고 있는 반면, 소수성 나노입자는 막에 직접 침투하여 이중층의 내부 소수성 코어에 매립되는 것으로 관찰되었습니다[139].
혼합 지질/고분자 막과의 QDNP 상호작용은 NP의 소수성 표면을 변화시켜 평가하였다. 소수성 NP는 막의 혼합 지질/고분자 단층에서 고분자 도메인 내에 위치하는 반면, 친수성 QDNP는 단층에 흡착되어 전체에 퍼져 공기/물 계면에서 분자 패킹에 더 높은 영향을 나타내는 것으로 관찰되었습니다[140] . 소수성 및 친수성 리간드가 혼합된 기능화된 AuNP를 리포솜 벽에 통합하는 것이 연구되었습니다. 결과는 소수성 리간드가 이중층의 소수성 코어와 상호작용하는 반면 친수성 리간드는 수용액과 상호작용한다는 것을 보여주었다[141].
표면 수정 효과
NP의 생의학 응용에서 NP의 표면 화학적 변형은 독성을 감소시키고 안정성을 증가시키며 NP의 세포 내재화를 제어하고 조절하여 생물학적 운명을 조절하는 데 사용되는 중요한 단계입니다[142]. NP의 표면 기능화는 주로 PEG, 음의 카르복실(-COOH) 기, 하이드록실(-OH) 기 같은 중성 관능기 및 포지티브 아민(-NH2) 기로 구성됩니다. (-NH2) 양의 증가는 표면 양전하를 증가시켜 NP가 세포로 흡수되는 것을 증가시킨다[143,144,145,146]. 유사하게, -COOH 작용기는 NP의 음전하를 증가시키고 그에 따라 흡수를 향상시킨다[144].
Tao et al. [147]은 종양 표적화를 위해 폴리도파민 기능화된 NP-압타머 생체접합체를 설계했습니다. 그들은 기능화된 NP가 비기능화된 NP에 비해 더 나은 표적화 효능을 가지고 있다고 보고했으며, 이는 향상된 치료 효과로 해석되는 기능화된 NP에 대한 더 높은 세포 흡수율을 나타냅니다. 또 다른 연구에서 엽산 기능이 있는 나노입자는 비기능화된 나노입자보다 자궁경부암 세포를 표적으로 삼는 데 더 높은 효능을 보였습니다[148]. AuNPs의 독성과 세포 흡수에 대한 표면 코팅의 영향은 Qiu와 동료들에 의해 연구되었습니다[90]. 그들은 폴리(디알릴디메틸 암모늄 클로라이드) 코팅된 AuNR이 세포에 의한 내재화에서 더 높은 효율을 보여주기 때문에 표면 코팅이 세포 흡수율을 결정하는 핵심 요소임을 밝혔습니다.
원래의 폴리스티렌(PS-NP)과 아미노 기능화된 폴리스티렌 NP의 세포 흡수 차이는 Jiang과 동료에 의해 조사되었습니다[149]. 결과는 아미노 기능화된 폴리스티렌 NP가 PS-NP보다 더 높은 흡수율을 가지며 전자는 주로 clathrin 매개 경로를 통해 내재화되고 후자는 clathrin-independent endocytosis를 통해 내재화된다는 것을 보여주었습니다. 이 놀라운 차이는 NP와의 세포 상호 작용에서 표면 화학적 변형의 핵심 역할을 강조합니다. 표면 개질 풀러렌, C60( C( 쿠)2)2 NP는 시간, 온도 및 에너지 의존적 방식으로 세포내이입을 통해 주로 세포에 의해 내재화되었습니다. C60의 내재화를 위해 선호되는 경로는 클라트린 매개 엔도사이토시스인 것으로 밝혀졌습니다. ( C( 쿠)2)2 NP [150].
탄력성 효과
NP 재생의 탄력성은 세포에 의한 내재화에 영향을 미치는 본질적인 요소입니다. NP의 탄성은 힘이 가해질 때 변화에 대한 저항으로 설명될 수 있습니다. 강성, 경도 및 강성은 NP의 탄성을 설명할 때 동의어인 용어 중 일부입니다. NP의 탄성을 측정하기 위해 사용되는 측정 지표는 영률(Young's modulus)이고 측정 단위는 파스칼(Pa)입니다. 이 측정을 기반으로 하여 Young's modulus 값이 높을수록 NPs 탄성이 높고 그 반대의 경우도 마찬가지입니다. NP에서 이 값을 측정하는 데 사용되는 분석 장치 또는 기기의 예로는 원자력 현미경, 레오미터 및 나노인덴터가 있습니다. 더 높은 탄성 값을 갖는 NP를 경질 NP라고 하며 이러한 NP의 예로는 금 NP, 양자점 및 자기 NP가 있습니다. 더 낮은 탄성 값을 갖는 NP를 연성 NP라고 하며, 이들의 예로는 하이드로겔, 리포솜 및 생분해성 고분자가 있습니다.
세포 흡수와 관련하여 NP의 이 매개변수에 초점을 맞춘 수많은 연구에서 부드러운 NP에 비해 더 뻣뻣한 NP를 더 효율적으로 내부화하는 세포의 선호에 대해 보고했습니다[151, 152]. 분명히, 이 관찰은 NP를 감싸는 데 필요한 변형 에너지가 내재화 과정 전반에 걸쳐 다양함에도 불구하고 더 부드러운 NP에 비해 더 단단한 NP를 감싸는 멤브레인에 의한 더 적은 전체 에너지 소비에 기인합니다. 또한, CGMD(Coarse-Grained Molecular Dynamics) 시뮬레이션을 사용하여 수행된 다양한 탄성을 갖는 NP의 멤브레인 랩핑에 대한 컴퓨터 모델링은 뻣뻣하고 부드러운 NP를 내부화하는 것과 관련된 변형 에너지 변화에 관한 실험적 관찰과 일치합니다[153]. However, there are also other studies that have reported on softer NPs being internalized more efficiently than stiffer NPs [154, 155] and intermediate elastic NPs internalized more efficiently compared to either stiff or soft NPs [156]. Hence, tuning the elasticity of NPs for better cellular internalization could be a valuable tool in biomedical applications such as drug delivery. A potential application was demonstrated by Guo and coworkers, whereby accumulation of nanolipogels in tumour cells were enhanced primarily by controlling this parameter of NP [157].
Intracellular Trafficking of NPs
In the previous sections, different possible uptake pathways of NPs and the parameters that affect the efficacy of uptake has been discussed. Following uptake, the next crucial matter is the intracellular trafficking of NPs which determines its final destination within cellular compartments, its cytotoxicity and its therapeutic efficacy [158, 159]. After NPs are internalized by the cells, they will first encounter membrane-bound intracellular vesicles called early endosomes. Endosomes formed at the plasma membrane are categorized into three types; early endosomes, late endosomes and recycling endosomes [106, 160,161,162,163].
Early endosome ferries the cargo to the desired cellular destination. Part of the cargo is recycled to the plasma membrane via recycling endosomes. Early endosomes transform into late endosomes via maturation and differentiation process. The late endosomes will then integrate with lysosomes to form endolysosomal vesicles and hydrolytic enzymes contained within these vesicles degrade the trapped NPs [18, 164,165,166]. However, some NPs are able to escape this pathway and are released into the cytoplasm therefore bypassing the lysosomal degradation process [167,168,169]. Another intracellular degradation pathway which plays important role in the intracellular fate of NPs is an intracellular process called autophagy [170,171,172]. In this process, cytoplasmic contents will be surrounded by autophagosome and delivered to the lysosome to be broken down and recycled [173]. In addition, aggregated proteins and dysfunctional organelles are degraded by autophagy to maintain cellular homeostasis. It is necessary to consider this pathway since recent studies demonstrated that several NPs are capable of inducing autophagy [174,175,176,177,178].
The intracellular trafficking of Tat peptide-conjugated quantum dots (Tat-QDs) in live cells was studied by Ruan and co-workers [179]. Dynamic confocal imaging showed that Tat-QDs interacted with negatively charged CMs leading to its internalization by macropinocytosis. The QD containing vesicles were observed to be actively transported by molecular motors towards the perinuclear region known as the microtubule-organizing center (MTOC). Tat-QDs bind to cellular membrane structures such as filopodia and vesicle shedding results in releasing QD-containing vesicles from the tips of filopodia.
The uptake and intracellular fate of fluorescent carboxylated polystyrene particles (20 nm and 200 nm in diameter) were evaluated by applying it on hepatocyte [180]. It was found that the particles were internalized by hepatocytes in size, time and serum-dependent manner. The fate of the particles was studied and they were not observed in early endosomes or lysosomes, but only in the mitochondria of the hepatocyte. Particles accumulated inside bile canaliculi show that NPs can be eliminated within bile. A study on the uptake and intracellular fate of silver NPs into human mesenchymal stem cells demonstrated that they agglomerate in the perinuclear region [181]. It was observed by using fluorescent probes that particles are contained within endo-lysosomal structures but not in the cell nucleus, endoplasmic reticulum or Golgi complex. Confocal imaging of FITC conjugated titania nanotubes in mouse neural stem cells revealed that they have crossed the karyotheca entering the cell nucleus [182]. Single-walled carbon nanotubes were observed to enter the cytoplasm and localize in the cell nucleus leading to cell mortality [183]. Translocation of AuNRs towards the nucleus has also been reported [184].
결론
The application of NPs in the modern world is growing at an exponential rate as the scientific enterprise is looking for novel ways to address current problems. NPs can be found as active ingredients in many formulations intended for human consumption, from cosmetics to processed foods. As its application increases in consumer products, so does human exposure to NPs. Hence, more research should be carried out to understand its potential hazards to humans and other living beings. In this review, we have looked at the current knowledge on the effects of NPs at a cellular level. Some of the topics discussed include cellular pathways of NPs and the influences of physiochemical properties of NPs on the uptake rate and uptake mechanism.