마그헤마이트 나노입자는 브라시카 나푸스의 성장 및 비생물적 스트레스 내성을 개선하는 나노자임으로 작용합니다.
초록
이트륨 도핑 안정화 γ-Fe2 O3 나노 입자는 식물 비료로 작용하고 효소 활동을 통해 가뭄 스트레스 관리를 지원하는 잠재력에 대해 연구되었습니다. 가뭄 후 과산화수소와 지질 과산화의 수준은 γ-Fe2일 때 감소했습니다. O3 나노 입자는 Brassica napus에 영양 용액의 관개에 의해 전달되었습니다. 토양에서 자라는 식물. 과산화수소가 151에서 83μM g
−1
으로 감소했습니다. 대조군과 비교하여 말론디알데히드 형성이 36에서 26mM g
−1
으로 감소했습니다. . 잎의 성장률은 완전히 수정된 식물에 비해 33%에서 50% 성장으로 향상되었으며 엽록소의 SPAD 측정값은 47에서 52로 증가하여 γ-Fe2를 사용하여 농경학적 특성이 개선되었음을 시사합니다. O3 킬레이트 철에 비해 비료로서의 나노 입자.
<섹션 데이터-제목="배경">
배경
식량 안보는 가장 중요하고 변화하는 세계의 시급한 문제입니다. 변화하는 기후와 증가하는 인구는 환경에 미치는 영향을 최소화하면서 식량 생산을 확보하기 위해 개선된 도구를 혁신하기 위해 식물 과학자와 농업 엔지니어를 이끌고 있습니다. 나노기술은 이러한 오랜 문제를 해결하기 위해 탐색할 수 있는 새로운 도구 중 하나입니다[1,2,3]. 나노기술은 예를 들어 작물 보호[4,5,6], 비료[7, 8], 바이오센서 및 정밀 농업[9]에서 역할을 하는 식품 생산 사슬의 중요하고 필수적인 부분이 될 것으로 예측되었습니다. 식품 포장 및 안전 [10]. 나노 입자는 자연에서 어디에나 존재하며 식물은 다양한 나노 입자에 노출되어 진화했습니다[11]. 산화철 나노입자(ION)는 자연적으로 발생하는 나노입자의 중요한 부분을 구성합니다[12]. 식물과 토양 미생물이 이온을 생성한다는 증거가 있습니다[11, 13, 14]. 일부 연구자들은 조작된 ION[15, 16]의 식물에 대한 독성에 대해 우려해 왔지만, 다른 연구자들은 ION을 비료로 사용할 가능성에 초점을 맞추었습니다[17,18,19,20,21,22]. 자철광 Fe3의 자성 나노입자 O4 및 마그헤마이트 γ-Fe2 O3 구조는 peroxidase 모방 능력(낮은 pH에서)과 카탈라아제 모방 능력(중성 pH에서) 모두의 효과적인 나노자임으로 제안되었습니다[23,24,25]. 특정 농도에서 나노 산화철은 킬레이트 형태의 동일한 양의 제1철 이온을 첨가하는 것과 비교하여 식물 성장을 증가시키는 것으로 나타났습니다[17]. 우리는 나노 산화철의 효소적 능력이 철 시비 이상의 식물 성장을 자극할 수 있다고 가정합니다. 또한, 우리는 이것이 카탈라아제와 퍼옥시다아제가 방출되는 활성 산소종(ROS)의 제거에 중요해지는 가뭄과 같은 일반적인 비생물적 스트레스 동안 식물을 도와야 한다고 제안합니다. 여기에서 우리는 γ-Fe2에 대한 이 가설을 테스트하기 위한 조사를 제시합니다. O3 토양과 통제된 환경에서 자란 유채 기름종자.
<섹션 데이터-제목="결과">
결과
식물 특성에 대한 입자의 영향
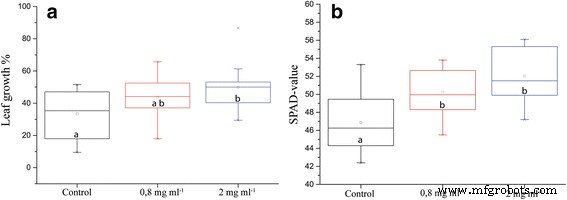
ION을 추가함으로써 우리는 적절한 양의 킬레이트화된 철을 추가하는 것에 비해 유지종자 평지의 성장을 증가시켰습니다. 잎 길이는 대조군과 비교하여 통계적으로 유의한 증가를 보여 세포 분열 또는 세포 신장의 증가를 시사합니다(그림 1a). 식물이 가뭄에 노출되기 전에 SPAD-미터로 측정한 엽록소 함량이 통계적으로 유의하게 증가했으며, 이는 대조군에 비해 이 식물의 적합성이 증가했음을 시사합니다(그림 1b).
<그림>
ION을 함유한 영양소 또는 영양소로 관개된 토양으로 화분에서 자란 식물의 다양한 생리학적 매개변수. 아 ION 처리 5일 후까지 개별 잎 길이 증가(n =16, p 값 =0.053). ㄴ SPAD 측정으로 측정한 잎의 엽록소 함량(n =16, p 값 =0.000). 다른 문자는 통계적으로 유의한 차이를 나타냅니다.
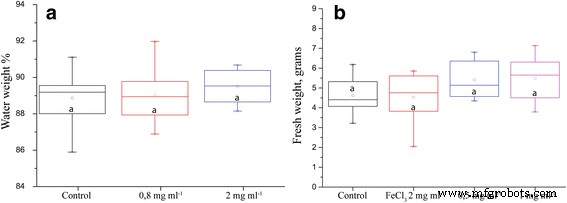
수분 손실은 통계적으로 유의미한 차이를 보이지 않았지만 ION 처리에서 수분 보유가 더 큰 경향이 있었습니다(그림 2a). 식물의 성장도 고려한 신선한 무게는 항상 ION 처리에 대해 더 높은 값을 보였고(그림 2b) 어떤 경우에는 통계적으로 유의했습니다. 예를 들어, 장기간 가뭄에 대한 한 실험을 그림 3에서 볼 수 있습니다.
<그림>
가뭄 스트레스 후 식물 매개변수. 아 물을 구성하는 식물 무게의 백분율. ㄴ 가뭄 5일 후 식물 바이오매스(n =8, p 값 =0.127). 다른 문자는 통계적으로 유의한 차이를 나타냅니다.
<그림>
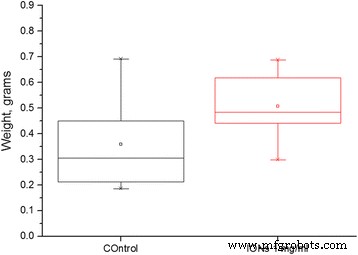
가뭄 5일 후 측정한 식물의 신선 중량입니다. 15개의 생물학적 복제 및 p로 통계적으로 유의한 차이 값 0.01
대조군에도 충분한 양의 철분이 함유되어 있다는 점을 고려할 때 신선 중량의 큰 차이는 이례적일 것입니다. ION 처리된 식물은 가뭄 기간 동안 대조군보다 더 잘 대처하고 물을 주면 더 잘 회복되는 것으로 관찰되었습니다(그림 4).
<그림>
가뭄 스트레스 5일 후 물을 뿌린 후의 식물 사진. 아 영양 용액으로 관개된 식물을 통제하십시오. ㄴ 0.8mg/ml 이온을 함유한 영양액으로 관개하는 식물. ㄷ 2mg/ml 이온을 함유한 영양 용액으로 관개된 식물
잎 과산화수소 농도에 대한 이온의 영향
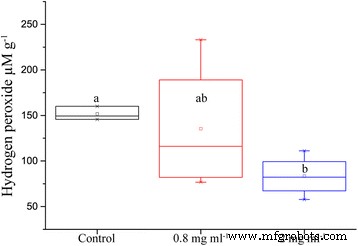
가뭄 후 잎에 있는 과산화수소의 양은 관수에 사용된 양액에 이온을 첨가했을 때 상당히 감소했습니다. 0.8mg ml
−1
에서 편차가 컸습니다. 치료; 따라서 다른 치료법과의 차이는 통계적으로 유의하지 않습니다. 그러나 대조군과 최고 농도인 2mg ml
−1
간의 차이는 p와 함께 통계적으로 유의미함 0.004의 값과 대조군 처리에서 평균 84% 더 큽니다(그림 5).
<그림>
ION을 함유한 영양 용액으로 처리하고 5일 동안 가뭄에 도전한 평지 잎 조직 그램당 가용성 과산화수소의 양(n =16, p 값 =0.004)
지질 과산화에 대한 ION의 효과
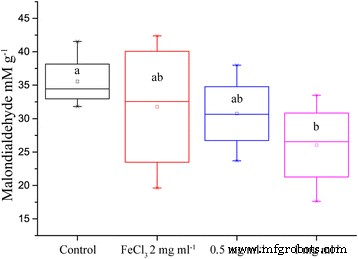
MDA 수준을 프록시로 사용하는 지질 과산화는 ION을 추가함으로써 감소되었으며, 200mg의 ION이 추가된 식물의 잎에서 MDA의 평균 농도는 36% 더 낮았습니다. 우리는 철(III) 이온의 몰 농도가 동일한 양성 대조군을 추가했습니다. 그러나 편차가 너무 커서 결론을 내릴 수 없었습니다. 낮은 ION 농도의 평균도 대조군보다 낮았으며, 이는 유채 잎에서 지질 과산화가 감소하는 경향을 보여줍니다(그림 6).
<그림>
5일 동안 ION 및 가뭄을 포함하는 양액으로 처리된 유채 잎에서 지질 과산화 생성물 MDA의 농도(n =8, p 값 =0.052)
식물 입자 흡수
잎 조직으로의 입자 흡수를 조사하기 위해 우리는 유도 결합 플라즈마 원자 방출 분광법(ICP-AES)으로 잎의 철 함량을 측정했습니다. 실제로, 철의 통계적으로 유의한 증가가 마그헤마이트 나노입자 처리에서 관찰되었습니다. 흥미롭게도, 철의 농도는 과잉 철(III) 이온으로 관개된 잎에서 감소했습니다(그림 7).
<그림>
마그헤마이트 나노입자 처리 후 브라시카 식물 잎의 철 농도는 철(III) 이온의 비율이 1:1M인 동일한 양액 또는 동일한 양액을 사용한 대조군과 비교했습니다. 다른 문자는 통계적으로 유의한 차이를 나타냅니다(n =15)
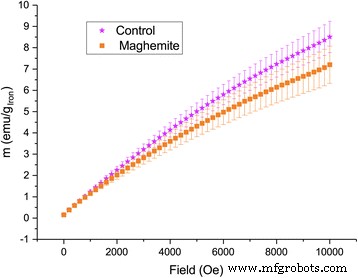
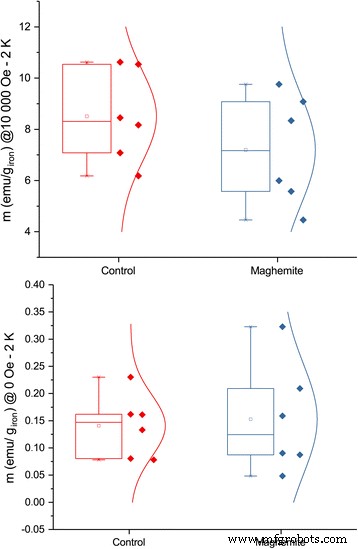
입자 흡수의 증거로서 증가된 철 함량을 더욱 확증하기 위해 동일한 잎에서 저온 자화를 측정했습니다. 강한 자기장 하에서 대조군에서 더 큰 자화가 관찰되었지만 자기장이 없을 때 잔류자화는 마그헤마이트 나노입자로 처리된 잎에서 더 컸다(그림 8 및 9). 작은 샘플 크기와 큰 변동으로 인해 그 차이는 통계적으로 유의하지 않지만 높은 자기장에서 제어에서 자화가 더 높지만 자기장이 없을 때 더 낮기 때문에 경향은 초상자성 ION의 존재를 명확하게 보여줍니다. 변동으로 인해 흐려지지만 특정 샘플에서는 ION의 존재가 명확하게 보였습니다(추가 파일 1:그림 S2). 한편으로, 충분히 낮은 온도와 충분히 높은 자기장에서 철 이온의 자화는 페리자성 이온의 자화보다 클 것입니다. 반면에 동일한 저온이지만 자기장이 0이면 나노입자 자기 모멘트가 차단되어 이온의 잔류 자화가 더 커질 것입니다.
<그림>
대조 식물과 비교하여 마그헤마이트 나노입자로 처리된 식물의 잿더미 잎의 저온(2 K) 자화. 오차 막대는 평균의 표준 오차를 보여줍니다(n =6)
<그림>
서로 다른 자기장 하에서 애쉬 잎의 저온(2K) 자화. 10000 Oe에서 위의 플롯에는 p가 있습니다. 값이 0.8이고 0 필드 아래의 플롯에는 p가 있습니다. 0.08의 값(n =6)
재료 특성
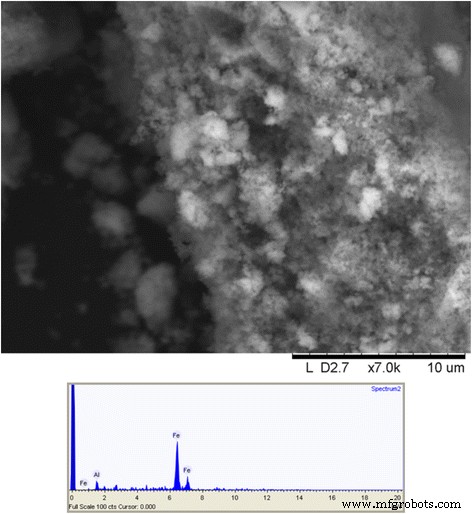
Cui et al.의 방법으로 생성된 ION. (2013) ~ 1–10s nm 정도의 나노입자가 성공적으로 생산되었음을 나타내는 겔을 형성했습니다. 건조된 겔을 분말로 분쇄하였다. 저해상도 SEM은 개별 입자를 표시할 수 없지만 분말의 계층 구조는 분명합니다. 시료의 EDS에서는 Y가 검출되지 않고 철만 검출되었습니다(그림 7).
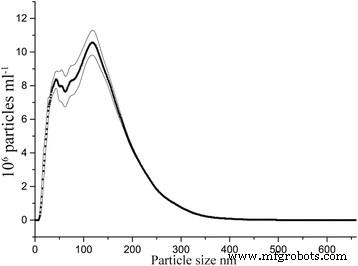
물에 분산되면 입자는 최대 500nm의 유체역학적 크기로 응집체를 형성하지만 응집된 입자의 84%는 300nm보다 작고 11% 이상은 50nm보다 작습니다. Nanosight 측정에 따르면 절대값은 4.28 × 10
6
입니다. 20nm ml보다 작은 입자
−1
, 측정에 필요한 50배 희석 분산액(그림 8). 다시 계산하면 약 2 × 10
8
이 있음을 의미합니다. 20nm ml보다 작은 입자
−1
치료 중입니다.
AFM으로 촬영한 이미지는 NTA와 XRD 비디오 인프라를 결합한 것과 유사한 패턴을 보여줍니다. 입자 크기는 수 나노미터에서 수백 나노미터까지입니다(그림 9).
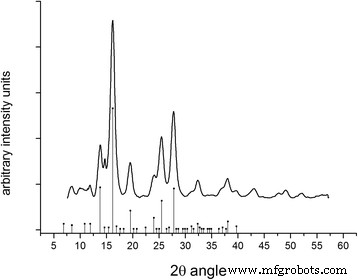
입자의 XRD는 생산 후 1년에 획득했으며 여전히 마그헤마이트 구조의 명확한 패턴을 보여 성공적인 마그헤마이트 안정화의 증거입니다(그림 10). 결정자 크기는 Scherrer 방정식을 사용하여 3.8nm로 계산되었습니다. 구조는 보존되지만 Y의 13%가 도입되면 물론 원자의 진동 상태에 영향을 미칩니다(추가 파일 1:그림 S3).
<그림>
γ-Fe2의 주사 전자 이미지 O3 이트륨 유도 졸-겔 합성 및 동일한 물질의 EDS 스펙트럼을 통해 합성
토론
철 비료로 ION의 제안된 사용은 다른 시스템에서 이전에 조사되었습니다[17,18,19]. 이번 조사에서는 중요한 작물에 미량 영양소인 철분을 공급하는 효과 외에 유사한 비료의 효소 효과가 있는지 여부를 처음으로 검증했다. 대조군에는 적절한 양의 킬레이트화된 철이 제공되었다. 우리는 또한 철(III) 이온의 몰당량을 식물에서 이용 가능한 적절한 양의 철로 음성 대조군에 보충한 양성 대조군을 테스트했습니다. 따라서 우리 실험에서 볼 수 있는 ION의 긍정적인 효과는 ION의 특성에서 비롯됩니다. 우리는 활동 중인 이온의 알려진 효소 효과라고 제안하고 싶습니다[23, 25, 26]. 다른 메커니즘은 여전히 배제할 수 없습니다. 이온은 또한 단백질, 지질 및 기타 생체분자와 상호작용할 수 있습니다[27], 또는 나노입자가 표면에 천연 철 이온을 흡수하여 유해한 Fenton 반응을 감소시킬 수 있습니다. ION 처리에서 잎의 과산화수소 수준이 감소했다는 사실 자체가 나노입자 흡수의 간접적인 증거입니다. 증가된 철 농도와 마그헤마이트의 변화된 자화와 함께 그림은 더욱 완벽해집니다. 철(III) 이온을 사용한 양성 대조군은 잎의 철 함량 농도가 감소했으며, 이는 식물이 방어 기제로 철 이온 흡수를 감소시키는 능력이 있음을 나타냅니다. 이것은 마그헤마이트 처리에서 잎에 있는 증가된 철 농도가 실제로 나노 입자이며, 이는 철 이온의 증가된 수준만큼 식물에 독성이 없음을 시사합니다. 자기 측정은 매우 작은 γ-Fe2의 전형적인 저온에서 초상자성 거동과 차단된 나노입자 자기 모멘트를 보여줍니다. O3 마그헤마이트로 처리된 잎에서 [28] 나노입자 흡수를 분명히 보여줍니다. 소량의 Y를 투여해도 식물에 영향을 미치지 않아야 합니다. 식물에 대한 Y의 영향에 대해서는 알려진 바가 거의 없지만 Fu et al. (2014) 2 mg L
−1
Y는 수경재배 시스템에서 치사량 중앙값(LD50)이었고 Maksimovic et al. (2014) 10
−5
에서 독성 효과를 보기 시작했습니다. 몰 L
−1
Y[29, 30]. 실험에 사용된 최고 농도에서 대략적인 양은 50mg(5.6 10
−4
mol)의 Y는 관개를 통해 포트당 토양에 첨가되었으며, 이로부터 일부만 흡수될 것으로 예상됩니다. 흡수되는 것은 이온으로 쉽게 이용 가능하지 않아야 하며, 오히려 마그헤마이트 입자에 결합되어야 합니다. 합성에 Y를 도입하는 목적은 마그헤마이트 나노입자의 용해도를 감소시키고 또한 철 산화물의 덜 효소적 형태인 적철광으로의 변형을 방지하는 것입니다. 의심할 여지 없이, 스트레스 조건 동안 카탈라아제 활성을 증가시키는 것이 유익합니다[31], 스트레스 조건의 전체 범위가 H2의 독성 축적을 유발하는 것으로 알려져 있기 때문입니다. O2 [32]. 또한 H2 O2 스트레스에 대한 신호 분자 역할도 합니다[32, 33]. 증가된 바이오매스 생산은 아직 확증되지 않았으며, 바람직하게는 유지종자 수확량과 품질도 조사하는 것이 좋습니다. 잎 성장 속도 증가, 잡초와의 경쟁이 중요한 농업 환경에서 매우 우수한 특성과 같은 다른 기능은 쉽게 고려할 수 있습니다. Arabidopsis에 표시되었습니다. 나노 0가 철 입자는 잎의 아포플라스트 내로 양성자의 압출을 유도하여 터거 구동 세포벽 확장을 가능하게 할 수 있습니다[34]. 동일한 효과가 뿌리에서도 관찰되었으며, 이는 가뭄 스트레스 동안에도 도움이 될 수 있습니다[35]. 그들은 또한 수분 손실로 이어질 수 있는 잎의 기공의 증가를 관찰했지만 측정했을 때 대조군과 약간의 차이만 있었습니다. 기공 개방과 수분 증발 사이의 관계가 선형이 아니라는 것은 알려진 역설입니다[36]. 이 관계는 또한 상대 습도나 바람과 같은 환경의 영향을 많이 받습니다[37]. 0가 나노입자는 물론 마그헤마이트와 동일한 것으로 간주되어서는 안 되지만, 우리 실험에서 볼 수 있는 잎 신장에 대한 메커니즘을 조사해야 합니다. Ghafariyat al. (2013)은 우리가 한 것처럼 철이 전혀 없는 음성 대조군과 비교하여 이온을 추가할 때 잎의 엽록소 농도가 증가하는 것을 관찰했습니다. 킬레이트 철과 비교할 때 차이가 없었습니다. 그러나 동일한 양의 킬레이트화 철을 이온으로 추가하면 철의 많은 부분이 결정 구조에 저장되기 때문에 식물에서 이용 가능한 철이 더 많아집니다. 따라서 식물은 실제로 철 결핍을 겪고 있는 이온으로만 수정되었을 가능성이 있습니다. 이온이 킬레이트화된 철에 보조적으로 추가되었을 때 잎에서 더 많은 양의 엽록소를 발견했습니다(SPAD 측정에 따르면, 그림 1 참조). 또한 가뭄 후 ION을 추가했을 때 잎에서 감소된 과산화수소와 MDA의 양을 측정했습니다. Rui et al. (2016)은 과산화수소를 측정하지 않고 산화 스트레스와 관련된 MDA와 효소를 측정했습니다. 그들은 산화 스트레스가 ION의 첨가로 인해 발생하지 않는다고 제안했으며 실제로 10mg kg
−1
에서 킬레이트화된 철에 비해 잎에서 감소된 MDA 양을 발견했습니다. 집중. 뿌리에서 그들은 ION의 농도를 증가시키면서 MDA의 감소를 보았습니다. 그들은 또한 킬레이트화된 철과 비교하여 감소된 수퍼옥사이드 디스뮤타제 및 퍼옥시다제 활성을 측정하여 ION이 생체 내에서 반응성 산소 제거제로 작용할 수 있다는 우리의 가설이 옳을 수 있음을 시사합니다. 활성 산소 제거는 Brassica napus 잎에서 측정된 과산화수소 감소에 의해 추가로 입증되었습니다. . 이것은 이온 첨가 시 관찰되는 가뭄에 대한 저항 증가를 설명합니다.
결론
우리의 실험은 식물체에서 나노자임으로 작용하는 ION의 메커니즘에 대한 증거를 제공했습니다. , Brassica napus 잎의 과산화수소 함량 감소 사이의 결합을 나타냄 및 ION의 도입. 따라서 이온 첨가 시 관찰되는 가뭄에 대한 저항 증가는 산화 스트레스 완화와 관련될 수 있습니다.
방법
실험 조건 및 디자인
브라시카 나푸스 봄 유채 품종 Larissa(Scandinavian Seed AB, Lidköping, Sweden)의 종자를 살균하고 3일 동안 한천 플레이트에서 발아시킨 후 유사한 크기의 묘목을 살균된 S-Soil(Hasselfors garden, Örebro, 스웨덴)이 있는 화분으로 옮겼습니다. . 모든 거대 및 미량 영양소, 통기용 펄라이트, 성장 촉진 부식산 및 pH 6이 적은 묘목의 전문적인 성장을 위한 토양입니다. 식물을 7일 동안 화분에 정착시키고 탈이온수로 관개했습니다. 물. 처리를 시작하기 전에 식물의 크기가 가능한 한 일정하도록 쟁반 사이에 식물을 분배했습니다. 7일째부터 화분으로 옮긴 후, 식물에 양액, 추가 FeCl이 첨가된 양액으로 관개3 또는 γ-Fe2 농도가 다른 영양 용액 O3 이온. 각 포트는 매일 40ml로 관개되었습니다. 식물은 16시간 빛(180μE m
-2
s
−1
) 및 8시간 어둠. 온도는 조사 시 25°C로 설정하고 어둠 속에서 22°로 설정하고 상대 습도를 65%로 설정했습니다. 식물은 각각 8개의 화분이 있는 트레이의 8 × 8 cm 화분에서 재배되었습니다. 모든 처리에는 2개의 트레이와 16개의 생물학적 복제물이 있었습니다. 트레이는 챔버의 변동을 보상하기 위해 매일 회전하는 순서로 이동되었습니다. 5일 동안 0.5, 0.8, 1 또는 2mg/ml의 총 200ml를 추가하여 치료를 계속했습니다.
−1
, 각각 식물당 총 100, 160, 200 또는 400mg입니다. ION을 추가한 5일 후, 모든 처리는 4일의 가뭄이 시작되기 전에 추가로 5일 동안 영양 용액(추가 파일 1:표 S1)으로 물을 주었습니다. 4일의 가뭄 후 과산화수소 및 지질 과산화 측정을 수행하고 식물에 3일 동안 동일한 양액으로 다시 물을 주어 회복을 연구했습니다. 실험은 4번 반복되었습니다.
나노입자 합성 및 특성화
약 13% 중량의 이트륨(Y)을 사용하여 [38]의 방법에 따라 마그헤마이트 입자를 제조하고 X선 회절(XRD), 주사 전자 현미경(SEM), 나노 입자 추적 분석(NTA), 적외선 분광기를 특징으로 합니다. (FTIR), 열중량 분석(TGA) 및 원자력 현미경(AFM). SEM 이미지는 Oxford μDeX 전자 분산형 X선 분광계(EDS)가 있는 Hitachi TM1000으로 획득했습니다. 유체역학적 크기는 Nanosight 300에서 NTA(나노 추적 분석)를 통해 측정되었습니다(그림 11). Perkin-Elmer Spectrum 100을 사용하여 브롬화칼륨(KBr) 펠릿에서 푸리에 변환 적외선 분광법(FTIR)을 수행했습니다. 열중량 분석(TGA)의 경우 Perkin-Elmer Pyris 1이 사용되었고 원자력 현미경(AFM)의 경우 Bruker FastScan이 사용되었습니다(그림 12). XRD는 몰리브덴 소스가 있는 Bruker Smart ApexII 다목적 회절계에서 수행되었습니다. 결정자 크기는 Origin 소프트웨어 피크 파인더 기능으로 계산된 1.01358489355378의 전체 너비 반값(FWHM)과 함께 2θ° 각도 16,197에서 최대 피크를 사용하여 셔러 방정식으로 계산되었습니다(그림 13). 건조된 ION은 3.4mg L
-1
의 영양 용액에 현탁되었습니다. 킬레이트 철, 대조군과 동일하게 사용됨. 모든 영양소의 전체 목록은 추가 파일 1:표 S1을 참조하세요.
<사진>
NTA로 측정한 γ-Fe2의 수중 유체역학적 입자 크기 분포 O3 이트륨 유도 졸-겔 합성을 통해 합성됩니다. 값은 4회 반복 측정의 평균이며 가는 선 안의 영역은 평균 오차를 나타냅니다.
<그림>
이트륨 유도 졸-겔을 통해 합성된 마그헤마이트 나노 입자는 실리콘 웨이퍼에 분산되고 AFM으로 이미지화됩니다. 동일한 이미지가 3D와 2D로 표현됩니다.
<그림>
생성된 마그헤마이트 나노입자의 분말 회절도. 피크는 그림의 베이스에 드롭 라인이 있는 점으로 본 데이터베이스의 표준 마그헤마이트 위치와 정렬됩니다. 결정 크기는 16.197 2θ에서 가장 큰 피크에서 계산되었습니다. 도 각도
식물 특성 측정
처리를 시작하기 전에 각 식물에서 가장 긴 잎인 첫 번째 실제 잎의 길이를 측정했습니다. 그 후, 5일간의 ION을 양액 또는 양액 단독으로 순차적으로 관수한 후 동일한 잎을 다시 측정했습니다. 결과는 백분율 증가로 보고됩니다. 잎 엽록소는 Minolta SPAD-meter로 SPAD 측정으로 처리 전과 처리 후, 가뭄 후에 평가되었습니다. 16개의 생물학적 복제물 각각에 대해 식물당 2개의 잎에 대한 3회의 측정값을 평균화했습니다. 마지막으로, 모든 식물의 지상 바이오매스의 무게를 재고 알루미늄 호일에 넣어 110°C에서 72시간 동안 건조했습니다.
철분 함량 및 자기 측정
5일간의 가뭄 후 실험이 종료되었고 지상의 모든 식물 조직은 450°C에서 24시간 동안 재처리되었습니다. 회분을 균질화한 후, 샘플당 10mg을 칭량하고 밤새 진탕기에서 36% 염산 3ml에 용해시켰다. 그런 다음 샘플을 Milli-Q 물에 녹인 10% 에탄올 44.74ml로 희석한 다음 238.204nm에서 ICP-AES로 철을 측정했습니다. 자기 측정을 위해 동일한 재를 샘플 홀더에 놓고 각 샘플의 정확한 무게를 정밀 저울로 칭량했습니다. 그런 다음 샘플을 2K로 냉각하고 SQUID(초전도 양자 간섭 장치) 자력계에서 10,000에서 0 Oersted로 자기장 스윕을 수행했습니다. 측정된 자기 모멘트에서 시료 홀더로 인한 자기 모멘트를 빼서 시료 내 철의 무게로 정규화합니다.
과산화수소 측정
잎의 과산화수소는 [39]에 의해 보고된 eFOX 방법으로 측정되었습니다. 16개의 식물을 각각 4개의 식물이 있는 4개의 생물학적 복제물로 풀링했습니다. 각 식물의 가장 젊고 아직 신선한 잎에서 50mg을 채취했습니다. 그런 다음 200mg의 잎 재료를 액체 질소로 미리 냉각된 모르타르에서 가루로 만들었습니다. 분말에 4ml의 100mM 인산 완충액(pH 6.9)을 추가하고 얼음을 균질한 액체로 막자사발에 담았습니다. 이 균질한 액체에서 1900μl를 2ml Eppendorf 튜브로 옮기고 20μl의 25mM ferrous ammonium sulphate(Mohrs salt), 20μl의 10mM 소르비톨, 20μl의 10mM 99 자일레놀 오렌지, 20μl를 추가했습니다. % 에탄올, 250mM 황산 20μl. 각 샘플에 대해 전체 가시광선 흡광도 스펙트럼을 사용했지만 550nm와 800nm의 차이를 과산화수소 정량에 사용했습니다. R을 사용하여 2~40μM의 과산화수소 보정 곡선을 만들었습니다.
2
0.9946의 값.
지질 과산화
지질 과산화는 [40]의 방법에 따라 측정하였다. 샘플을 4ml 0.1% w로 균질화한다는 점을 제외하고는 과산화수소 측정과 동일한 방식으로 채취했습니다. /v 트리클로로아세트산(TCA). 532nm에서 흡광도를 측정하고 600nm에서 흡광도를 빼서 비특이적 탁도를 보정했습니다. 155mM cm
−1
의 소광 계수 말론디알데히드 농도(MDA)를 계산하는 데 사용되었습니다.
통계 분석
모든 통계는 Minitab 17 소프트웨어에서 수행되었습니다. 모든 데이터는 그룹화를 위해 Fisher 테스트를 사용한 일원 분산 분석을 통해 실행되었습니다. 학생의 t 특정 p를 찾기 위해 테스트를 수행했습니다. 통계적으로 유의한 차이가 있는 것으로 밝혀진 그룹 간의 값.