알츠하이머병(AD)은 노년층에서 가장 흔한 신경퇴행성 질환입니다. 알츠하이머병의 치료에서는 BBB(Blood-Brain Barrier)를 통한 약물 침투의 어려움, Aβ 펩타이드의 불충분한 제거, 염증 인자의 대량 방출과 같은 몇 가지 장애물을 시급히 극복해야 합니다. 이러한 문제를 해결하기 위해 우리는 BBB를 통한 약물의 침투를 향상시키기 위해 키토산(CS)과 소 혈청 알부민(BSA)으로 구성된 특수하고 새로운 나노 입자(NP)를 개발했습니다. 강력한 항염증제인 커큐민을 사용하여 Aβ 펩타이드의 식균 작용을 증가시켰습니다. 결과는 curcumin-loaded CS-BSA NP가 BBB를 통한 약물 침투를 효과적으로 증가시키고 미세아교세포의 활성화를 촉진하며 Aβ 펩티드의 식균 작용을 더욱 가속화한다는 것을 보여주었습니다. 또한, curcumin-loaded CS-BSA NP는 TLR4-MAPK/NF-κB 신호 전달 경로를 억제하고 M1 대식세포 분극을 더욱 하향조절했습니다. 이 연구는 커큐민이 로딩된 CS-BSA NP가 알츠하이머병에서 대식세포 분극을 조절함으로써 Aβ 42 식균 작용을 향상시킬 가능성을 시사했습니다.
소개
알츠하이머병(AD)은 교활한 발병과 인지 쇠퇴의 진행을 특징으로 하는 신경 퇴행성 장애입니다. 조직병리학적으로 42개의 아미노산을 포함하는 펩타이드인 아밀로이드-β(Aβ) 42는 알츠하이머병에서 세포외 "노인성 플라크"를 특징으로 하는 아밀로이드 β-펩타이드 원섬유로 응집되어 신경 세포 사멸과 시냅스 손실을 유발합니다[1,2,3] . 일반적으로 Aβ 42 단량체는 생리학적으로 가용성이고 독성이 없는 반면, 올리고머는 시험관 내 및 생체 내에서 더 독성이 있습니다[4, 5]. 따라서 Aβ 42 단량체를 제거하여 Aβ 42 응집을 중재하는 것이 알츠하이머병에 대한 가장 적절한 치료 표적으로 널리 간주됩니다[6,7,8,9]. Aβ 펩타이드는 TLR4를 비롯한 여러 Toll-like 수용체(TLR)와 상호작용하여 미세아교세포 활성화를 유발할 수 있으며, CD14, TLR4 또는 TLR2 의존성 식세포 작용과 Aβ42 제거를 촉진한다는 것은 잘 알려져 있습니다[10, 11,12]. 소교세포 활성화는 Aβ 42의 제거를 촉진할 수 있지만, 단핵 대식세포의 한 유형인 소교세포는 과활성화되어 M1 표현형으로 분극화될 수 있습니다(잠재적으로 신경독성 가용성 인자 및 전염증성 사이토카인의 방출을 특징으로 함). 신경 세포의 죽음과 AD의 발병을 악화시킵니다. 반대로, 일부 소교세포는 항염증성 사이토카인의 생산을 특징으로 하는 고전적으로 활성화된(M2) 표현형입니다. 이들은 AD의 인지 기능 장애를 개선합니다[13,14,15,16]. 따라서 M1/M2형 대식세포의 비율은 알츠하이머병의 진행에 상당한 영향을 미칠 수 있습니다[17, 18]. 또한, 전염증성 M1을 항염증성 M2 대식세포로 전환하는 데 필요한 상향 조절은 알츠하이머병 예방 및 치료에 유망한 잠재력을 보여줄 것입니다.
생강의 일종인 인도 향신료 심황(Curcumin longa)의 성분에서 유래한 커큐민은 염증을 감소시킬 수 있고 알츠하이머병 치료에 중요한 역할을 할 수 있는 강력한 항염증제입니다[19]. 최근 몇 년 동안, 커큐민은 잠재적인 신경 보호 효과가 있을 수 있는 항아밀로이드 생성, 항염증, 항산화 및 금속 킬레이트 특성을 가지고 있는 것으로 보고되었습니다[20, 21]. 커큐민은 toll-like receptor 4-mitogen-activated protein kinase(TLR4-MAPK)/NF-κB 경로의 억제를 통해 대식세포 분극을 조절합니다[22,23,24]. 그러나 curcumin의 열악한 안정성과 생체 이용률은 임상 적용을 제한합니다. 또한 BBB(혈액뇌장벽)의 존재는 알츠하이머병 치료에서 커큐민의 침투를 방지합니다[25,26,27].
혈액에서 뇌로의 약물 수송을 향상시키기 위해 펩타이드[28]와 항체[29]로 표면 기능화된 나노입자(NP)는 BBB를 통한 약물 전달을 돕고 NP의 BBB 침투 효율은 다른 능동 수송 메커니즘을 통해 크게 향상될 수 있습니다. 단순 수동 확산보다 [30]. 키토산(CS) 나노입자(Aβ와 결합하기 위한 나노운반체)는 BBB를 투과할 수 있으며 면역원성이 없습니다[31]. 또한 혈청 알부민은 인체의 순환 혈장에서 혈청 50g/L 농도로 발견되었으며 무독성이며 면역계에 잘 견딘다[32, 33]. 또한 소혈청알부민(bovine serum albumin, BSA) 유래 나노입자는 약물의 반감기를 증가시켜 투여 빈도를 감소시키고 환자의 순응도를 높일 수 있는 지속 방출 특성을 갖고 있다고 보고되었다[34]. 따라서 우리는 최고의 BBB 침투를 달성하기 위해 curcumin이로드 된 CS-BSA NP를 준비하기 위해 두 가지 생체 재료로 CS와 BSA를 사용했습니다. Aβ 42의 식균 작용, 염증성 사이토카인 분비 및 TLR4-MAPK/NF-κB 경로의 조절에 대한 커큐민의 효과를 조사하여 대식세포 분극에 대한 커큐민의 분자 메커니즘을 추가로 확인했습니다.
자료
탈아세틸화도가 80%이고 분자량이 약 400kDa인 CS는 Haixin Biological Product Co., Ltd.(중화인민공화국 닝보)에서 구입했습니다. BSA는 Sigma-Aldrich Co.(미국 미주리주 세인트루이스)에서 구입했으며 커큐민은 Dalian Meilun Biotechnology Co., Ltd.(중화인민공화국 다롄)에서 구입했습니다. FITC-β-아밀로이드(1-42)는 Chinese Peptide Co., Ltd.(중국 항저우)에서 구입했습니다. 구매한 다른 화학물질은 분석 등급으로 Sigma-Aldrich Co.에서 구입했습니다. 대식세포 세포주 RAW 264.7(마우스 백혈병 단핵구 대식세포 세포주) 및 모델 역할을 하는 뇌 미세혈관 내피 세포주(hCMEC/D3) 인간 BBB의 세포 생물학 상하이 연구소, 중국 과학 아카데미, 상하이(중화 인민 공화국)에 의해 설립되었습니다. 두 전지 모두 37°C의 온도와 5% CO2에서 유지되었습니다. 대기, 10%(부피/부피) 열 불활성화 소태아 혈청 및 항생제(100U/mL의 페니실린 및 100mg/mL의 스트렙토마이신)로 보충된 Dulbecco의 변형 독수리 배지(DMEM)에서. 지질다당류(LPS)로 처리된 RAW 264.7 세포는 알츠하이머병으로 예시되는 신경퇴행성 질환에서 관찰되는 미세아교세포의 양상을 요약하는 것으로 보고되었습니다[35, 36]. 따라서, 지질다당류(LPS, 1μg/ml)에 의해 M1 표현형으로 극성화된 대식세포 세포주 RAW 264.7 세포를 추가로 적용하여 AD의 미세아교 세포를 시뮬레이션했습니다.
커큐민 함유 CS-BSA NP 준비
우리의 이전 보고서[37]에 따르면, 정전기 상호작용 하에서 양전하를 띤 CS는 음전하를 띤 BSA와 결합하여 NP를 형성할 수 있습니다. 제조 방법은 다음과 같았습니다:0.1% 아세트산을 사용하여 CS를 용해하여 CS 0.5mg/mL의 용액을 얻고 0.05mg/mL의 커큐민을 포함하는 DMSO 100μL를 CS 용액에 첨가하여 자기 하에서 완전히 혼합했습니다. 실온에서 교반. 적절한 양의 BSA 용액으로 1.0 mg/mL를 CS와 커큐민의 혼합물에 천천히 적가했습니다. 이 시점에서 유백광 현상이 나타났고 CS-BSA NP는 고체 입자로 더 응축되었다. 또한 NP의 크기, 다분산성, 제타 전위 및 형태를 조사했습니다. 나노입자로부터의 시험관내 약물 방출은 이전에 보고된 방법을 사용하여 추정되었습니다[37]. NP에서 커큐민의 캡슐화 효율(EE, %)은 아래 식을 사용하여 계산되었습니다.
세포 사멸에 대한 CS-BSA NP의 안전성을 결정하기 위해 MTT 분석을 사용하여 세포 생존력을 평가했습니다. 이전 연구의 프로토콜에 따르면 추가 분석을 위해 다양한 양의 빈 CS-BSA NP를 사용하여 RAW 264.7 세포(M1 표현형) 및 hCMEC/D3 세포를 37°C에서 24시간 동안 처리했습니다.
체외 BBB 모델을 사용한 침투 연구
뇌 미세혈관 내피 세포주인 hCMEC/D3를 사용하는 단층 트랜스웰 배양은 NP의 뇌 전달을 연구하는 데 사용되는 일반적인 시험관 내 BBB 모델입니다. hCMEC/D3 세포 믹스(총 부피 0.5–1.0mL)를 300Ω 이상의 경내피 전기 저항을 갖는 단층 세포 배양을 위한 12웰 트랜스웰 플레이트의 상부 챔버에 삽입물에 추가했습니다. pH 7.4의 PBS를 하부 챔버에 첨가하였다. 유리 커큐민과 커큐민이 로드된 CS-BSA NP의 현탁액을 37°C, 5% CO2로 설정된 배양기에서 3시간 동안 연속 배양을 위해 상부 챔버에 넣었습니다. . 그 후, 유리 커큐민과 커큐민이 로딩된 CS-BSA NP가 세포를 가로질러 옮겨져 하부 챔버에 들어갔습니다. 침투된 NP의 정량은 425nm에서 여기되고 530nm에서 방출되는 커큐민의 형광 강도를 확인하여 마이크로플레이트 리더(Synergy-2; BioTek Instruments, Winooski, VT, USA)를 사용하여 감지되었습니다. 나노입자의 투과율을 나타내는 상대형광비(RFR, %)는 초기에 첨가된 커큐민의 형광강도에 대한 하부 챔버에 침투된 커큐민이 로딩된 CS-BSA 나노입자의 형광강도의 비율을 결정하여 계산하였다. 상부 챔버에 CS-BSA NP를 로드했습니다. 10μg/mL의 클로르프로마진(클라트린 매개 흡수 억제), 1μg/mL의 제니스테인(카베올라 매개 흡수), 사이토칼라신 D(30μM, 거대음세포증), 20μg/mL의 나트륨과 같은 다양한 세포내이입 억제제 azide(에너지 억제제)는 다양한 침투 메커니즘과 관련된 다양한 세포내이입 경로를 확인하는 데 사용되었습니다. 상대 침투율은 억제제로 처리된 NP의 침투율과 비억제제를 처리한 NP의 침투율을 비교하여 결정되었습니다.
커큐민이 함유된 CS-BSA NP의 세포 흡수
RAW 264.7 세포(M1 표현형)에서 커큐민이 로딩된 CS-BSA NP의 분포와 위치는 공초점 레이저 스캐닝 현미경(FluoView FV10i; Olympus Corporation, Tokyo, Japan)을 사용하여 관찰되었습니다. hCMEC/D3 세포 믹스(총 부피 0.5–1.0mL)를 300Ω 이상의 경내피 전기 저항을 갖는 단층 세포 배양을 위한 12웰 Transwell 플레이트의 상부 챔버에 있는 삽입물에 추가했습니다. RAW 264.7 세포(M1 표현형)를 하부 챔버에 접종했습니다. 유리 커큐민 및 커큐민 로딩 CS-BSA NP의 현탁액을 37°C, 5% CO2로 설정된 인큐베이터에서 연속 배양을 위해 상부 챔버에 넣었습니다. . 공초점 레이저 스캐닝 현미경을 사용하여 커큐민이 방출하는 녹색 형광을 감지하여 RAW 264.7 세포(M1 표현형)에서 자유 커큐민 및 커큐민이 로드된 CS-BSA NP의 세포 분포를 미리 결정된 간격으로 관찰했습니다.
유리 커큐민 및 커큐민 부하 CS-BSA NP에 의해 유도된 Aβ 42의 식균 작용 감지
완전 성장 배지의 RAW 264.7 세포(M1 표현형)를 12웰 플레이트(1 × 10
5
세포/웰) 및 37°C에서 24시간 동안 유리 커큐민 및 커큐민 로딩 CS-BSA NP로 처리했습니다. 내부화되지 않은 curcumin과 curcumin이 포함된 CS-BSA NP를 제거하기 위해 증류수를 사용하여 단시간에 2번 세포를 세척했습니다. curcumin 및 curcumin-loaded CS-BSA NP는 배지에서 완전히 제거되었으며, 증류수로 세척된 세포의 형태가 손상되지 않고 세포 파열이 없기 때문에 증류수로 인한 세포가 낮은 삼투압을 유도할 명백한 위험이 없음이 발견되었습니다. 관찰되었다. 마지막으로 PBS(pH 7.4)에 용해된 유리 FITC 표지 Aβ 42를 플레이트에 첨가하여 3시간 동안 연속 배양했습니다. RAW 264.7 세포(M1 표현형)로의 FITC-표지된 Aβ42의 식균작용은 FITC에 의해 방출되는 녹색 형광을 검출함으로써 나타내었다. 공초점 레이저 스캐닝 현미경(FluoView FV10i; Olympus Corporation)을 사용하여 세포 내 식균 작용 및 세포 내 Aβ 42의 위치를 추가로 연구했습니다.
서부 얼룩 분석
커큐민 매개 대식세포 분극의 가능한 분자 메커니즘을 탐구하기 위해 우리는 종양 괴사 인자(TNF)-α 및 인터루킨(IL)-6과 같은 전염증성 사이토카인의 발현 수준과 ERK, JNK의 인산화 수준을 조사했습니다. , p38 및 NF-κB를 웨스턴 블롯팅하여 TLR4-MAPK/NF-κB 신호 전달 경로에 대한 커큐민의 추가 연구 특이적 효과
<섹션 데이터-제목="결과">
결과
커큐민이 포함된 CS-BSA NP의 특성
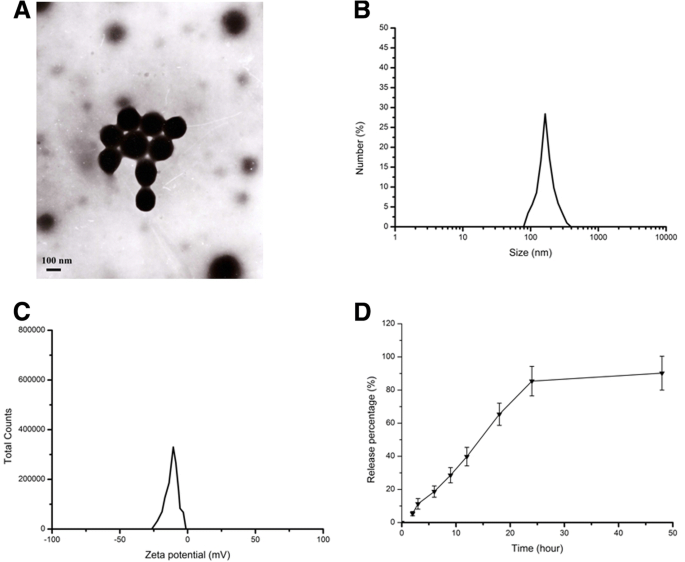
나노입자의 입자크기, 제타전위 및 형태를 결정하기 위해 나노입자의 가속전압에서 Zetasizer(Nano ZS90; Malvern Instruments, Malvern, UK)와 투과전자현미경(TEM)(Jeol, Tokyo, Japan)을 이용하여 나노입자의 특성을 조사하였다. 200kV. 그림 1에 표시된 결과는 CS-BSA NP가 각각 143.5nm에서 평균 크기, - 10.8mV에서 음의 제타 전위, 0.021에서 다분산성을 나타냄을 나타냅니다. curcumin-loaded CS-BSA NP는 모양이 구형이고 단분산된 것으로 관찰되었습니다. curcumin-loaded CS-BSA NPs를 포함하는 생성된 현탁액을 원심분리하여 상등액을 얻어 curcumin의 흡광도를 측정하고 표준 곡선에 따라 상등액에서 유리 curcumin의 함량을 계산했습니다. NP에서 커큐민의 캡슐화 효율(EE, %)은 95.4%로 평가되었습니다. 나노입자로부터의 약물 방출 과정의 관점에서, 커큐민이 로딩된 CS-BSA 나노입자는 pH 수준이 7.4인 배지에서 2상 방출 패턴을 보였다. 모든 약물의 약 11.3%가 처음 3시간 이내에 방출되었는데, 이는 NP가 BBB에 도달하기 전에 혈액 순환에 들어갔을 때 커큐민이 잘 보호되고 NP의 코어에 캡슐화되었음을 나타냅니다. 또한 몇 가지 약물이 NP에서 누출되어 처음 3시간 이내에 혈액으로 방출되었습니다. 대부분의 curcumin-loaded CS-BSA NP는 BBB 주위로 수송될 수 있고 뇌 주위의 약물 농도를 향상시킬 수 있습니다.
<그림>
커큐민 부하 CS-BSA NP의 특성화. 아 커큐민 로딩 CS-BSA NP의 TEM 이미지. ㄴ 획득한 커큐민 부하 CS-BSA NP의 동적 광산란(DLS) 분석. ㄷ 얻은 커큐민 부하 CS-BSA NP의 제타 전위 분석. d 48시간 동안 37°C에서 pH 7.4인 인산완충식염수에서 얻은 커큐민 로딩 CS-BSA NP의 시험관 내 방출 프로필
체외 BBB 모델을 사용한 침투 연구
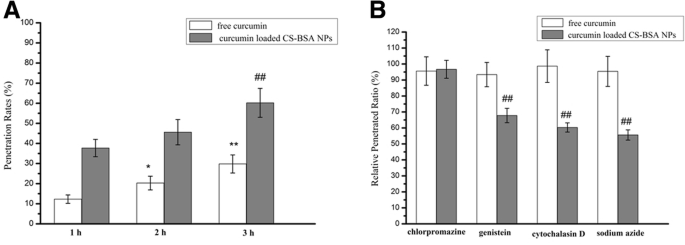
마이크로플레이트 리더(Synergy-2; BioTek Instruments)를 이용하여 하부 챔버의 커큐민 형광강도를 확인하여 유리 커큐민과 나노입자의 투과율을 평가하였고, 투과된 커큐민의 형광강도의 비율을 구하여 계산하였다. -하부 챔버에 로딩된 CS-BSA NP를 상부 챔버에 초기에 추가된 커큐민 로딩 CS-BSA NP의 그것으로. 결과(그림 2)는 유리 커큐민과 커큐민이 포함된 CS-BSA 나노입자의 침투 과정이 시간에 따른 패턴을 따르고 시간이 지남에 따라 침투율이 증가함을 보여주었습니다. 이는 유리 커큐민의 침투율이 1시간에 12.3%, 2시간에 20.3%, 3시간에 29.8%임을 시사했습니다. 유리 커큐민과 비교하여 커큐민이 함유된 CS-BSA 나노입자의 침투 효율은 증가된 침투율로 표시되는 것처럼 향상되었습니다. 침투율은 1시간에 37.7%, 2시간에 45.6%, 3시간에 60.2%로 증가했습니다. 이것은 유리 커큐민이 세포를 통해 침투하는 데 어려움을 겪을 수 있고 BBB에 대한 낮은 투과성을 보일 수 있음을 나타냅니다[38, 39]. 이 관찰은 또한 curcumin-loaded CS-BSA NP가 세포를 통한 약물 침투를 효과적으로 촉진할 수 있음을 나타내어 다양한 세포내이입 경로가 수행하는 역할을 시사합니다. 엔도사이토시스 억제 시험은 이전 보고서[40]와 일관되게 유리 커큐민은 침투를 위한 수동 확산에 의존하며 억제제의 추가 여부와 상관없이 유리 커큐민의 침투 효율에 명백한 변화가 없음을 보여주었습니다. 반대로 나노입자의 침투는 에너지 의존적이었고, 아지드화나트륨으로 처리한 상대 침투율은 55.6%였다. 또한, 동굴과 거대음세포증은 주로 NP의 세포내이입 경로를 매개합니다. 비억제제 처리와 비교하여 제니스테인과 시토칼라신 D 처리한 세포의 상대 침투율은 각각 67.8%와 60.3%였습니다.
<그림>
hCMEC/D3 세포를 통한 자유 커큐민 및 커큐민 부하 CS-BSA NP의 침투 메커니즘 분석. 아 자유 커큐민과 커큐민이 포함된 CS-BSA NP의 투과율에 대한 형광 스펙트럼 분석. 결과는 평균 ± 표준편차(n =3). *피 <0.05, **P <0.01 대 1시간에 유리 커큐민의 침투율.
##피 <0.01 대 1시간에 커큐민 부하 CS-BSA NP의 침투율. ㄴ 자유 curcumin 및 curcumin-loaded CS-BSA NP의 침투 능력에 대한 endocytic inhibitors의 효과. 결과는 평균 ± 표준편차(n =3).
##피 <0.01 대 클로르프로마진으로 처리된 커큐민 부하 CS-BSA NP의 상대 침투 비율
MTT에 의한 세포 사멸 평가
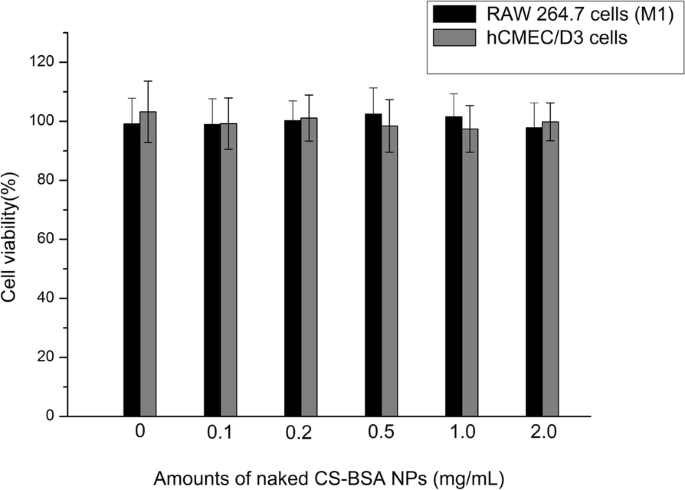
RAW 264.7 세포(M1) 및 hCMEC/D3에 대한 블랭크 CS-BSA NP의 세포독성 효과는 MTT 분석에 의해 시험관내에서 추정되었다. 세포를 0~2.0mg/mL 범위의 다양한 농도의 CS-BSA NP로 처리했습니다. 그림 3의 세포 생존력 분석은 빈 CS-BSA NP로 처리했을 때 RAW 264.7 세포와 hCMEC/D3 세포에서 명백한 세포독성 활성이 관찰되지 않았음을 보여주었습니다[37].
<그림>
24시간(n =3)
NP의 분포 및 세포 흡수
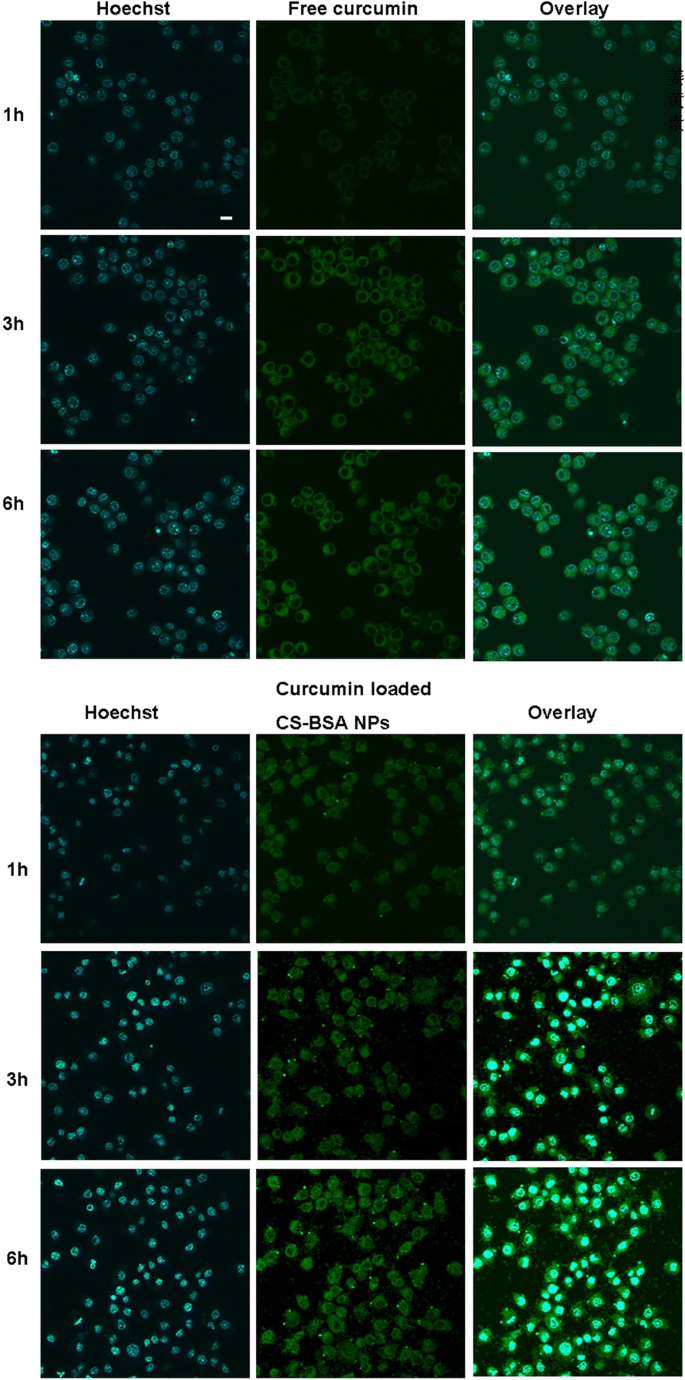
100μg/mL의 동일한 양의 커큐민을 함유하는 자유 커큐민 및 커큐민 로딩 CS-BSA NP를 사용하여 RAW 264.7 세포(M1)를 처리했으며, 공초점 레이저 스캐닝 현미경(FluoView FV10i)으로 커큐민의 분포 및 세포 흡수를 관찰했습니다.; 올림푸스). 그림 4에서 유리 커큐민과 커큐민이 로딩된 CS-BSA 나노입자의 세포 내 분포가 시간 의존적 패턴을 따르고 커큐민의 녹색 형광이 세포 내에 축적되어 전체 세포질에 분산되었음을 알 수 있습니다. 시간이 지남에 따라 세포 내부의 녹색 형광 강도가 향상되었습니다. 이것은 세포 내부의 유리 커큐민이 방출하는 녹색 형광이 매우 약하다는 것을 보여주었으며, 이는 대부분의 유리 커큐민이 대식세포 세포주인 RAW 264.7에 의해 흡수되지 않았음을 나타냅니다. NP는 매우 효율적인 세포 내 약물 전달을 위한 유망한 잠재력을 가질 수 있기 때문에 [41, 42], curcumin-loaded CS-BSA NP는 유리 curcumin으로 처리된 세포와 비교할 때 증가된 형광 강도를 나타내는 것으로 관찰되었으며, 이는 CS-BSA NP가 개선될 수 있음을 시사합니다. 커큐민의 세포 흡수. 여기에서 이 관찰은 CS-BSA NP가 세포 내에서 약물 축적을 효과적으로 촉진할 수 있음을 나타냅니다.
<그림>
6시간 동안 RAW 264.7 세포(M1)에서 자유 커큐민 및 커큐민 로드 CS-BSA NP의 흡수. 커큐민은 녹색 형광색을 나타내었고 자유 커큐민과 커큐민이 로딩된 CS-BSA NP의 세포 내 위치를 나타냈습니다. 핵은 37°C에서 15분 동안 Hoechst(파란색)로 염색되었습니다. 스케일 바는 50μm이며 모든 그림 부분에 적용됩니다.
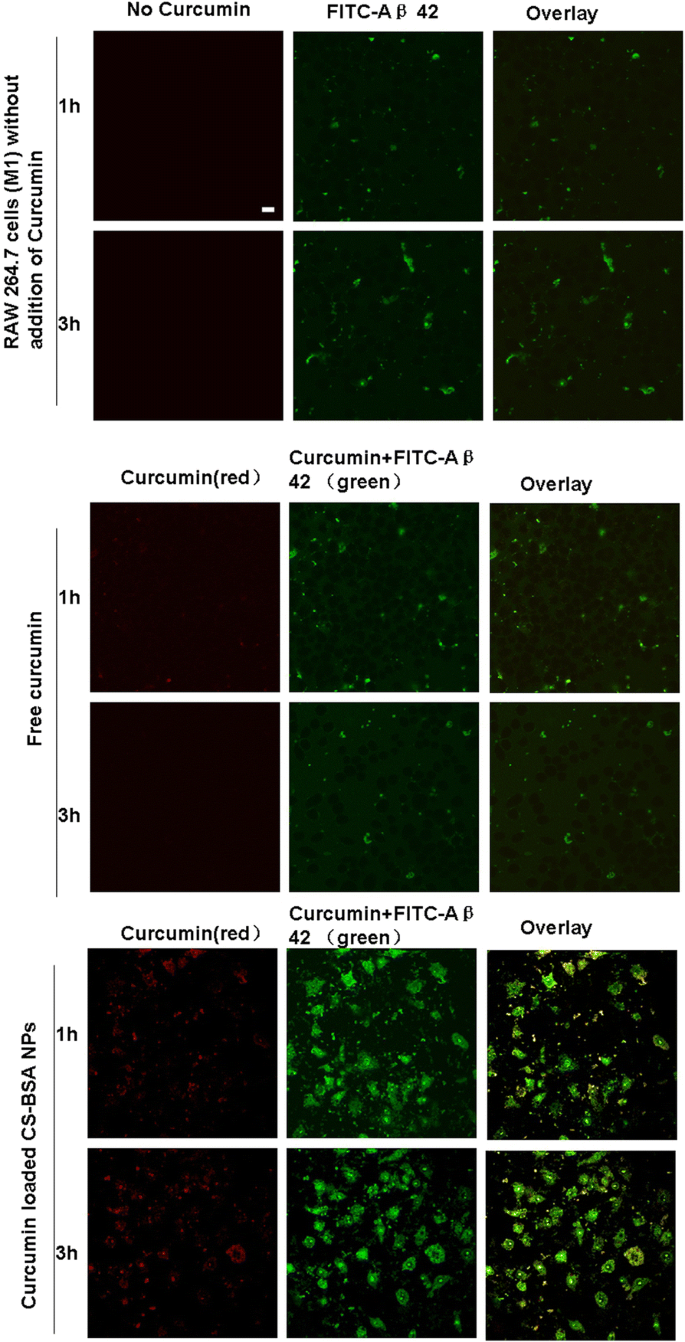
유리 커큐민 및 커큐민 부하 CS-BSA NP에 의해 유도된 Aβ 42의 식균 작용
도 5에 나타난 바와 같이 커큐민은 여기 파장 550nm, 방출 파장 570nm에서 적색 형광을 나타냈다. 또한, FITC의 여기 및 방출 파장에서도 녹색 형광을 보였다. 따라서 curcumin 및 FITC로 표지된 Aβ 42가 RAW 264.7 세포(M1)로 식균되면 모두 FITC의 여기 및 방출 파장에서 녹색 형광을 나타냅니다. co-localization 실험에서 적색 형광과 녹색 형광(커큐민을 나타냄)이 병합되었고 노란색 점은 커큐민의 세포 내 존재와 위치를 나타냅니다. 일부 녹색 점은 RAW 264.7 세포(M1)에서 FITC로 표지된 Aβ 42의 존재 및 식균 작용을 나타내는 적색 형광 점과 같은 위치에 있지 않았습니다. 적은 양의 Aβ 42가 미세아교세포에 의해 식균되는 것으로 관찰되었으며[43], 이는 세포에서 녹색 형광의 세포내 관찰에 의해 입증되었습니다. 그림 5의 오버레이 이미지에서 볼 수 있듯이 유리 커큐민에 비해 커큐민의 노란색 형광 점이 세포 내부에 더 많이 축적되어 노란색 형광으로 표시되는 curcumin이 로딩된 CS-BSA NP가 다량으로 존재함을 시사합니다. 강도는 NP와 RAW 264.7 세포(M1) 사이의 상호 작용으로 인해 세포에 축적되었습니다. 이것은 더 높은 세포내 농도의 커큐민을 유도하여 더 높은 녹색 형광 강도로 표시되는 Aβ 42의 식세포 작용을 증가시킵니다[44]. 커큐민이 포함된 CS-BSA NP는 대식세포 분극을 유도할 뿐만 아니라 항염증 및 신경 보호 효과가 식균 작용 강화에 기여한다고 가정했습니다.
<그림>
유리 커큐민 및 커큐민 부하 CS-BSA NP에 의해 유도된 Aβ 42의 식균 작용. 스케일 바는 50μm이며 모든 그림 부분에 적용됩니다.
서부 얼룩 분석
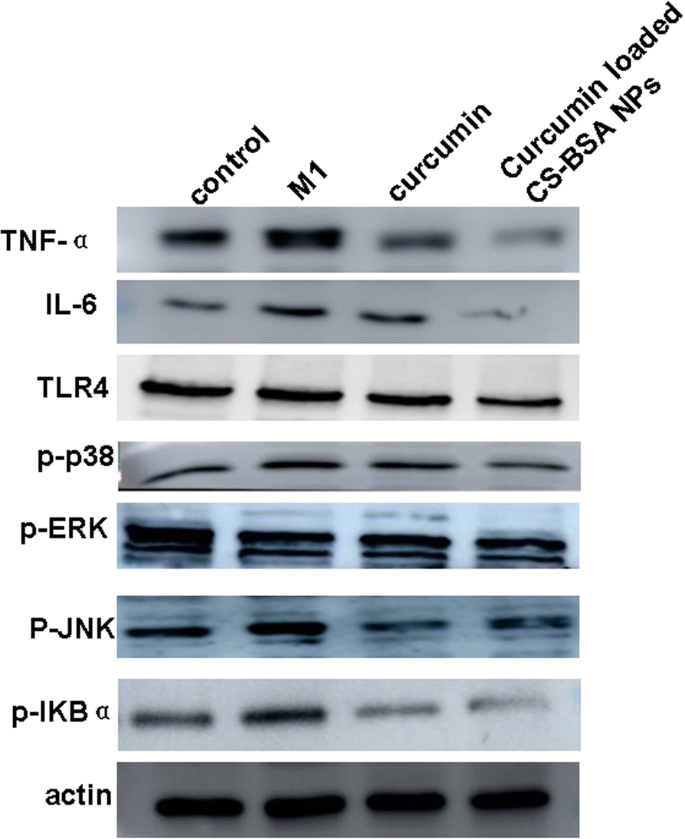
커큐민 매개 대식세포 분극의 가능한 분자 메커니즘을 탐구하기 위해 우리는 TNF-α, IL-6 및 TLR4의 전체 수준과 p38, ERK, JNK 및 IκBα의 인산화 수준을 웨스턴 블로팅으로 조사하여 추가 연구를 수행했습니다. TLR4-MAPK/NF-κB 신호 전달 경로에 대한 커큐민의 특정 효과
그림 6은 대조군인 정상 RAW 264.7 세포와 비교할 때 RAW 264.7 세포(M1 표현형)가 더 높은 수준의 TNF-α 및 IL-6 발현을 특징으로 하는 잠재적인 신경독성 가용성 인자 및 전염증성 사이토카인을 더 많이 방출한다는 것을 보여주었습니다. 따라서 신경 세포의 죽음을 초래하고 알츠하이머병의 발병을 악화시킵니다. 대조군 및 유리 커큐민과 비교하여, 커큐민이 로딩된 CS-BSA NP는 M1 대식세포 분극을 억제하고 RAW 264.7 세포(M1 표현형)에서 TNF-α 및 IL-6의 가장 낮은 발현을 유도하는 것으로 나타났습니다. 또한, 커큐민이 로딩된 CS-BSA NP는 M1 대식세포 분극화와 ERK, JNK, p38 및 NF-κB의 인산화를 조절하는 TLR4 발현을 감소시키는 것으로 나타났습니다. 이는 커큐민이 로딩된 CS-BSA NP가 세포 내에서 커큐민 축적과 이에 따른 세포 내 농도를 효과적으로 촉진하여 TLR4-MAPK/NF-κB 신호 전달 경로에 대한 차단 효과를 강화하고 M1 대식세포 분극을 더욱 억제한다는 것을 시사합니다. <그림>
유리 커큐민 및 커큐민 처리 후 RAW 264.7 세포(M1 표현형)에서 TNF-α, IL-6, TLR4의 발현 수준 및 ERK, JNK, p38 및 핵 인자(NF)-κB의 인산화에 대한 웨스턴 블롯 분석 로드된 CS-BSA NP
토론
알츠하이머병은 가장 흔한 신경퇴행성 질환 중 하나이며 선진국에서 주요 사망 원인입니다. 알츠하이머병의 치료와 관련하여 BBB의 뇌로의 대부분의 외인성 물질의 진입을 막는 능력은 사용을 막는 주요 장애물이었습니다. 이 문제를 해결하기 위해 BBB를 통한 커큐민의 수송을 개선하기 위해 CS-BSA NP를 설계했습니다. 결과는 이전 연구[45]와 일관되게 유리 커큐민이 세포를 통과하기 어렵고 BBB를 통과하게 되어 투과 효과가 더 낮다는 것을 입증했습니다. Curcumin-loaded CS-BSA NPs는 caveolae 및 micropinocytosis 매개 경로를 매개로 BBB를 통한 약물 침투를 효과적으로 촉진할 수 있습니다. Aβ 응집에 의해 유발된 신경염증은 알츠하이머병의 병리학적 기전의 기초가 되는 중요한 요인 중 하나입니다[46]. 뇌의 Aβ 수준은 Aβ의 생성과 제거 사이의 동적 균형에 의해 결정됩니다. 따라서 Aβ의 제거는 뇌에서 Aβ의 수준을 결정하는 데에도 중요합니다. 미세아교세포의 식균작용 능력은 알츠하이머병 예방에 중요한 생리학적 중요성을 나타냈다. 미세아교세포는 주로 Aβ-표적 제거를 담당하고 Aβ 42의 침착 영역 주변에서 주로 응집되는 경향이 있어 식세포 작용에 의한 Aβ 42의 축적을 추가로 방지합니다[47, 48]. 결과는 Aβ 42에 대한 커큐민 부하 CS-BSA NP 처리 RAW 264.7 세포(M1 표현형)의 식균 작용 효과가 증가하고 Aβ 42의 축적 및 침착이 감소하여 알츠하이머병 발병을 개선할 수 있음을 보여주었습니다.
미세아교세포(M1 표현형)는 잠재적으로 신경독성 가용성 인자와 TNF-α 및 IL-6과 같은 전염증성 사이토카인을 방출하여 신경세포 사멸을 유발하고 알츠하이머병 발병을 악화시킵니다[49]. M1 대식세포 분극은 TLR4-MAPK /NF-κB 신호전달 경로의 활성화에 의존하고 TLR4-MAPK /NF-κB 신호전달 경로를 차단하면 M1 대식세포 분극을 억제하고 M1 유형에서 대식세포의 분극을 촉진할 수 있다는 것이 발견되었습니다 M2 유형 [50]. 우리의 결과는 생강의 일종인 인도 향신료인 강황(Curcumin longa)의 성분인 커큐민이 염증을 감소시킬 수 있고 알츠하이머병 치료에 역할을 할 수도 있는 강력한 항염증제임을 보여주었습니다. CS-BSA NP는 효율적인 BBB 표적 커큐민 침투를 위한 강력한 도구 역할을 했습니다. 유리 커큐민과 비교할 때, 커큐민이 로딩된 CS-BSA NP는 식균 작용을 유도했으며 더 많은 양의 Aβ 42가 RAW 264.7 세포에 의해 식균되었습니다. 또한 curcumin이 포함된 CS-BSA NP는 유리 curcumin보다 TNF-α, IL-6 및 TLR4의 낮은 단백질 발현 수준을 유도했으며 ERK, JNK, p38 및 NF-κB의 인산화도 억제되었습니다. 커큐민이 로딩된 CS-BSA NP는 TLR4-MAPK/NF-κB 신호 전달 경로를 차단하여 M1 대식세포 분극을 억제하여 커큐민의 항염증 및 신경 보호 효과를 촉진함으로써 커큐민 유도 대식세포 식균 작용을 향상시킬 수 있음을 나타냅니다.
결론
우리의 데이터는 커큐민이 알츠하이머병 치료에서 치료제로 사용될 수 있음을 시사했습니다. 커큐민이 로딩된 CS-BSA NP는 커큐민의 향상된 BBB 침투 및 더 높은 세포내 약물 농도에 의해 Aβ 42의 RAW 264.7 세포 유도 식균 작용을 촉발했습니다. 또한, 커큐민이 로딩된 CS-BSA NP는 M1 대식세포 분극을 억제하고 TLR4-MAPK/NF-κB 신호 전달 경로를 차단하여 항염증 및 신경 보호 효과를 유도했습니다. 종합하면 커큐민이 함유된 CS-BSA NP는 AD 치료를 향상시키는 잠재력을 보여주었습니다.