F-액틴은 MKL1 및 YAP/TAZ를 통해 TiO2 나노튜브에서 중간엽 줄기 세포의 조골 세포 분화를 조절합니다
초록
티타늄과 티타늄 합금은 정형외과 임플란트에 널리 사용됩니다. 나노토포그래피를 수정하는 것은 티타늄 기판의 골유착을 개선하기 위한 새로운 전략을 제공합니다. 필라멘트 액틴(F-액틴) 중합은 기계적 로딩 구조로서 일반적으로 세포 이동, 세포 내이입, 세포 분열 및 세포 모양 유지에 관여하는 것으로 간주됩니다. F-액틴이 관여하는지 여부와 그것이 나노튜브로 유도된 중간엽 줄기 세포(MSC)의 골형성 분화에서 어떻게 기능하는지는 아직 밝혀지지 않았습니다. 이 연구에서 우리는 TiO2를 제작했습니다. 양극 산화에 의해 티타늄 기판 표면에 나노튜브를 만들고 주사 전자 현미경(SEM), X선 에너지 분산 분석(EDS) 및 원자간력 현미경(AFM)으로 그 특징을 특성화했습니다. Alkaline phosphatase (ALP) 염색, Western blotting, qRT-PCR, immunofluorescence 염색을 통해 골형성 가능성, F-actin 수치, MKL1과 YAP/TAZ 발현을 알아보았습니다. 우리의 결과는 TiO2의 내경과 거칠기 나노튜브는 양극 산화 전압이 30V에서 70V로 증가함에 따라 증가했으며 높이는 2μm 일관되게 유지되었습니다. 또한, 튜브 직경이 클수록 TiO2의 능력이 강해집니다. 나노튜브는 MSC의 골형성 분화를 촉진합니다. Cyto D에 의한 F-액틴 중합 억제는 MSC의 골형성 분화와 빈쿨린(VCL) 및 국소 접착 키나제(FAK)와 같은 국소 접착 복합체에 포함된 단백질의 발현을 억제했습니다. 대조적으로, Jasp 처리 후, F-액틴의 중합은 RhoA 및 전사 인자 YAP/TAZ의 발현을 증가시켰다. 이 데이터를 기반으로 TiO2 나노튜브는 MSC의 골형성 분화를 촉진했으며, 이 능력은 특정 범위(30-70V) 내에서 나노튜브의 직경이 증가함에 따라 향상되었습니다. F-액틴은 MKL1 및 YAP/TAZ를 통해 이 과정을 중재합니다.
소개
티타늄과 티타늄 합금은 생체 적합성, 내식성 및 기계적 특성이 우수하여 전체 관절 교체 및 치과 임플란트와 같은 임상 응용 분야에서 널리 사용됩니다[1,2,3]. 그러나 무균성 이완 및 감염 등 아직 해결해야 할 과제가 많이 남아 있다[4, 5]. 최근에는 골유착 및 항균성을 향상시키기 위한 많은 연구가 수행되고 있다. 예를 들어 MoS2 티타늄 임플란트의 /PDA-RGD 코팅은 티타늄 임플란트와 호스트 뼈의 통합을 촉진할 뿐만 아니라 고효율로 박테리아 성장을 억제할 수 있습니다[6]. 또한, 표면 지형은 점점 더 많은 주목을 받고 있으며 지형 변형은 마이크로 및 나노 규모의 구조만 변경함으로써 화학적 변형과 다릅니다. 세포에 대한 화학 신호의 자극은 불안정하고 세포 독성이 있습니다. 이에 비해 안전하고 제어 가능한 물리적 신호는 화학 분자로 인한 부작용을 피할 수 있습니다. 따라서 임플란트 표면의 지형적 변형과 지형적 구조를 통한 골유착 조절은 보철물 식립 후 골유착 불량이라는 임상적 문제를 해결하는 새로운 방법을 제시할 수 있다.
골 조직 공학 및 골 재생 분야에서 세포-형태 상호 작용은 종자 세포 기능 및 분화의 정확한 제어를 위한 유망한 관리 전략으로 간주됩니다. 동시에 뼈 자체는 나노미터와 마이크론 범위 내에서 우아한 계층 구조를 가지고 있습니다[7]. 따라서 표면 형태는 자연적인 뼈 구조를 모방하고 숙주 뼈와 임플란트의 표면에서 중간엽 줄기 세포의 골 형성 분화를 촉진할 수 있는 유사한 틈새를 제공할 수 있습니다. 표면 형태는 나노튜브, 나노와이어, 나노포어 등을 포함한 다양한 구조로 구성될 수 있습니다. 특히, 나노튜브 어레이는 높은 표면 대 부피비, 생물학적 가소성 및 높은 흡착 능력과 같은 독특한 표면 특성으로 인해 최근 여러 분야에서 많은 관심을 받고 있습니다. 예를 들어, 새로운 연구에 따르면 질화붕소 나노튜브(BNNT)는 오일에 용해된 가스의 조성과 함량을 감지하여 변압기의 작동을 모니터링하는 가스 센서로 사용할 수 있는 가스에 민감한 물질을 구성합니다[8]. 생물 의학에서 표면 지형은 세포 이동, 접착, 증식 및 분화를 포함한 세포 행동을 지시할 수도 있습니다. 최신 연구에 따르면 나노 규모의 지형은 중간엽 줄기 세포(MSC)가 조골 세포로 분화하도록 지시하여 조기 골유착을 강화할 수 있음을 보여줍니다[9,10,11,12]. 마이크로 및 나노 스케일의 결합된 표면 변형으로 MSC가 수축성 평활근 세포로 분화될 수 있다는 보고도 있습니다[13]. 그러나 표면 지형이 세포 운명을 어떻게 지시하는지에 대한 분자 메커니즘은 아직 해명되지 않았으며 이는 재료 안전성 평가 및 재료 설계에 중요합니다.
마이크로필라멘트라고도 하는 사상(F)-액틴은 진핵 세포에서 세포 골격의 세 가지 주요 구성 요소 중 하나입니다. 그것은 수많은 다른 단백질에 의해 변형된 구형(G)-액틴의 중합체로 구성됩니다. F-액틴은 모든 마이크로필라멘트의 소단위체가 같은 끝을 향하고 있기 때문에 구조적 극성을 가지고 있습니다. 미늘 끝은 다른 인접한 단량체로 향하고 뾰족한 끝은 ATP 결합 부위가 노출된 액틴 소단위를 가지고 있습니다. 즉, ATP는 G-액틴과 F-액틴 사이의 변환 과정에 관여합니다. 이 과정은 동적 평형 상태에 있으며, 중합과 해중합이 동시에 발생하며, 트레드밀이라고도 하며 라멜리포디아와 필로포디아에서 흔히 볼 수 있습니다[14]. 따라서 액틴 역학은 세포 이동, 세포 분열 및 세포 모양 유지와 같은 세포 기능에서 중요한 역할을 한다는 것은 분명합니다. 그러나 F-액틴은 기계적 부하를 지원하는 물리적 구조로 작용할 뿐만 아니라 신호 전달 및 유전자 발현과 같은 다른 생물학적 행동에도 참여합니다. 축적된 증거는 F-액틴이 다른 단백질과 상호작용하여 물리적 신호를 화학적 신호로 변환할 수 있음을 보여줍니다[15,16,17,18]. 예를 들어, 생체역학 및 기하학적 재구성은 F-액틴에 대한 액틴 단량체 중합의 중첩을 방지함으로써 종양 세포의 세포자멸사를 촉진합니다[15]. 초음파 펄스는 F-액틴의 해중합을 억제하여 인간 중간엽 줄기세포의 골형성을 향상시킵니다[16]. 우리의 이전 연구는 기계적 변형이 F-액틴의 안정성을 증가시키는 것으로 나타났습니다[17]. 나노튜브의 속이 빈 구조는 세포에 대한 접착 부위가 적기 때문에 생체역학적 균형을 유지하기 위해 세포골격의 재배열이 불가피합니다. 결과적으로, 우리는 F-액틴이 나노토포그래피에 의해 유도된 세포 분화를 매개할 가능성이 있다고 믿을 만한 이유가 있습니다.
이 연구에서 우리는 TiO2를 제작했습니다. 나노튜브는 양극 산화에 의해 지형을 수정하고 MSC의 골 형성 분화를 촉진하는 능력을 탐구했습니다. 다음으로, 우리는 F-액틴이 기계적 변환에서 중요한 역할을 하는지 여부를 조사했습니다. F-actin의 미늘 말단에 경쟁적으로 결합하여 G-actin이 필라멘트에 혼입되는 것을 방지하는 Cytochalasin D(Cyto D)를 사용하여 F-actin 중합을 억제하고 jasplakinolide(Jasp)를 사용하여 필라멘트의 안정화를 향상시켰다. 액틴 어셈블리. 또한, 우리는 F-액틴이 물리적 신호를 생화학적 신호로 바꾸는 데 어떻게 기능하는지 설명하고 싶었습니다. 이전 연구의 결과를 바탕으로 우리는 MAPK 경로가 이 과정에 포함될 수 있다고 가정했습니다[17]. Mechanosensors 및 mechanotransducers로 간주되는 PDZ-binding motif(TAZ) 및 MKL1을 포함하는 Yes-associated protein(YAP)/transcriptional coactivator와 같은 전사 인자도 F-actin이 줄기 세포에 미치는 영향을 선별하기 위한 본 연구의 대상이었습니다. 다른 분야의 일부 연구에서는 F-actin과 관련이 있다고 암시했기 때문입니다[19,20,21]. 일반적으로 우리는 나노튜브에 의해 유도되는 줄기세포 분화 과정에서 F-액틴의 역할을 명확히 하여 나노튜브로 변형된 임플란트의 재료 설계 및 생물학적 안전성 평가를 안내하기를 희망합니다.
자료 및 방법
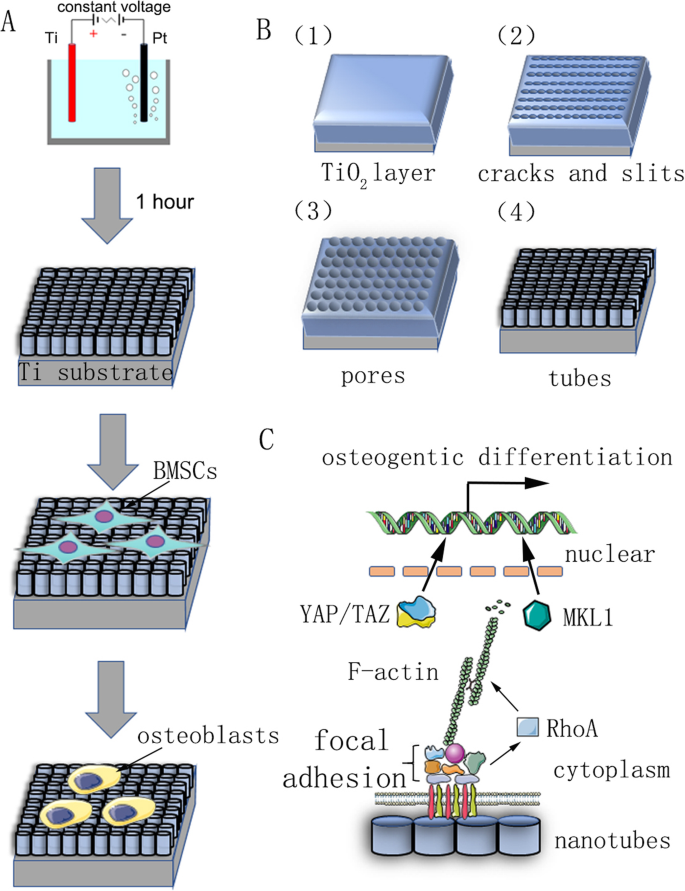
TiO 제작2 나노튜브
기판으로 사용된 순수 티타늄 조각(순도 99.9%, 두께 2mm; Shengshida, Hebei, China)은 400번 및 1500번 그릿의 탄화규소 사포로 연마되었습니다. 그 후, 샘플을 초음파 세정기에서 아세톤, 무수알코올, 탈이온수로 차례로 세척하고 최종적으로 상온에서 3시간 동안 건조시켰다. 나노토포그래피를 제작하기 위해 전처리된 샘플을 양극으로 고정하고 백금 조각을 0.15M NH4 전해질 수용액에서 반대 음극으로 사용했습니다. 1시간 동안 F 및 90% 글리콜. 양극 산화 전압은 30, 40, 50, 60, 70 V의 정전압이었다. 양극 산화 후 모든 시료는 탈이온수로 30분간 헹구고 초음파 세척기에서 무수알코올로 15분간 세척하였다. 마지막으로 모든 샘플을 120°C의 오토클레이브에서 1시간 동안 멸균한 후 배양액에 적셔 사용했습니다.
나노튜브 제작의 반응 메커니즘은 명확하지 않으며 현재 주류 이론은 현장 강화 용해 이론입니다. 나노튜브 어레이의 형성은 필드 산화, 필드 용해 및 화학적 용해의 작용 하에서 동적 평형의 결과입니다(그림 1b). 양극 산화 공정은 다음과 같이 설명할 수 있습니다. 첫 번째 단계에서 산화물 장벽 층이 전해질-금속 계면에 형성됩니다.
우리 연구의 요약 차트. 아 양극 산화 및 세포 분화 유도를 보여주는 흐름도. ㄴ 전처리된 티타늄 기판은 0.15M NH4가 포함된 전해질 수용액에서 양극으로 고정되었습니다. 1시간 동안 일정한 전압에서 F 및 90% 글리콜. 자기조립된 나노튜브는 균일하게 형성될 것으로 예측되었다. 그 반응 메커니즘은 재료 및 방법에 설명되어 있습니다. ㄷ 나노튜브에 의해 유도된 줄기세포의 골형성 분화 메커니즘의 개략도
그런 다음 산화물 층의 전계 강화 용해로 인해 표면에 균열과 좁은 슬릿이 나타납니다. F
−
의 확산 이온이 이러한 균열과 슬릿으로 들어가면 용해 속도가 향상됩니다. 균열은 확대되어 인접한 균열과 연결됩니다. 마지막으로 산화티타늄 층의 형성 속도와 용해 속도가 동적 균형에 도달하고 나노튜브가 더 이상 성장하지 않습니다.
다른 전압(30, 40, 50, 60, 70 V)으로 제작된 샘플을 에탄올과 탈이온수로 15분 동안 헹구고 실온에서 건조시켰다. 주사 전자 현미경(SEM450, FEI Nova Nano SEM; Thermo Fisher Scientific, Waltham, MA, USA)을 사용하여 표면 구조를 특성화하고 샘플을 얇은 금 층으로 코팅한 후 나노튜브의 내부 직경과 높이를 측정했습니다. 한편, 나노튜브의 원소 조성을 분석하기 위해 X선 에너지 분산 분석(EDS)을 수행하였다. AFM(Atomic force microscopy)(AFM, NanoManVS, Bruker Nano Surfaces, Bruker MicroCT, Kontich, Belgium)을 사용하여 샘플의 표면 형태와 표면 거칠기를 조사했습니다. 각 샘플에서 세 가지 다른 영역을 선택하고 측정을 세 번 반복했습니다.
세포 배양
4주 된 수컷 Sprague-Dawley(SD) 쥐는 Shanghai Ninth People's Hospital(Shanghai, China)의 실험 동물 센터에서 구입했습니다. 쥐의 골수 중간엽 줄기 세포(BMSC)는 대퇴골과 경골에서 무균적으로 분리되었습니다. BMSC를 정제하고 10%(v/v) 태아 소 혈청(FBS)을 포함하는 α-최소 필수 배지(α-MEM; Hyclone, Logan, UT, USA)(Gibco/Life Technologies, Carlsbad, CA, USA)에서 확장했습니다. ), 100 mg/mL 스트렙토마이신(Gibco) 및 100 U/mL 페니실린(Gibco)을 사용하고 95% 공기와 5% CO2로 구성된 습한 분위기에서 37°C에서 배양합니다. . 2일마다 배지를 교체하고 세포를 트립신 처리하고 80% confluence에서 계대배양하였다. 이 연구에 사용된 모든 세포는 계대 3과 5 사이였습니다. 골형성 유도 배지는 100nM 덱사메타손, 10mM β-글리세로포스페이트 및 50mM 아스코르브산이 보충된 성장 배지로 구성되었습니다(Sigma-Aldrich, St Louis, MO, USA). .
세포 증식 분석
TiO2 나노튜브로 변형된 티타늄 조각을 원형으로 자르고 24웰 세포 배양 플레이트의 웰에 넣었습니다. 계대 3-5 사이의 BMSC는 TiO2에서 배양되었습니다. 3 × 10
4
밀도의 나노튜브 성장 배지 또는 골형성 배지에 있는 세포/디스크. 세포 배양 2일 후, F-actin 중합을 방해하는 데 사용되는 cytochalasin D(Cyto D, Sigma-Aldrich)와 jasplakinolide(Jasp, Sigma-Aldrich)를 3일 동안 매일 배지에 첨가하였다. Cyto D와 Jasp의 최종 농도와 작업 시간은 각각 5μM, 1시간 및 2μM, 3시간이었습니다. 시약과 함께 인큐베이션한 후 배양 배지를 갱신했습니다. 세포 생존 및 증식은 Cyto D 또는 Jasp 처리 12시간 후 Cell Counting Kit-8(CCK8) 분석(Dojindo, Kumamoto, Japan)을 사용하여 평가되었습니다. 세포를 5% CO2하에 37°C의 세포 배양기에서 2시간 동안 10%(v/v) CCK8 용액과 함께 배양했습니다. . 그런 다음 100μL의 반응 혼합물을 96웰 플레이트의 웰에 옮기고 Multiscan UV-visible spectrophotometer(Safire2; TECAN, Mannedorf, Switzerland)를 사용하여 배양액에서 formazan 염료 생성물의 흡광도(OD)를 450nm에서 측정했습니다. ). 또한 자동 세포 계수기(AMQAX1000, Life Technologies)를 사용하여 대략적인 세포 수를 계산했습니다. 세포 계수 전에 BMSC는 TiO2에서 효소적으로 분리되었습니다. 나노튜브 및 Trypan blue(Sigma–Aldrich)로 염색되었습니다.
알칼리성 인산분해효소 염색 및 ALP 활성 분석
BMSC는 5개의 다른 TiO2에 시딩되었습니다. 3 × 10
4
밀도의 나노튜브 수정 티타늄 조각(30, 40, 50, 60 또는 70V) 웰당 및 골형성 배지에서 배양된다. 시약을 위에서 설명한 대로 추가했습니다. 배양 7일 후, TiO2에서 배양된 BMSC 나노튜브를 PBS로 3회 세척하고, 4% 파라포름알데히드로 고정하고, 제조업체의 지침(Hongqiao, Shanghai, China)에 따라 ALP 키트의 알칼리성 포스파타제(ALP) 작업 용액에서 인큐베이션했습니다. 결과는 PBS로 세척한 후 실체현미경으로 관찰하였다.
ALP 활성 분석을 위해 먼저 프로테아제 및 포스파타제 억제제가 없는 RIPA 완충액으로 세포를 용해한 다음, 제공된 프로토콜에 따라 ALP 분석 키트(Beyotime Institute of Biotechnology, Jiangsu, China)를 사용하여 원심분리된 용해물을 분석했습니다. 활성은 최종적으로 해당 용해물의 단백질 농도로 정규화되었습니다.
면역세포화학
Cyto D 및 Jasp 처리 3일 후, BMSC를 실온에서 20분 동안 4% 파라포름알데히드로 고정한 다음 PBS로 3회 세척하였다. 세포를 0.3% Triton-X 100으로 30분 동안 투과화하고 PBS로 3회 세척하고 어두운 곳에서 실온에서 1시간 동안 Rhodamine-conjugated phalloidin으로 염색하였다. 그런 다음 세포를 PBS로 헹구고 실온에서 10분 동안 DAPI(Beyotime Institute of Biotechnology)로 대조염색했습니다. PBS로 3회 추가 세척한 후 샘플을 유리 슬라이드에 고정하고 공초점 현미경으로 관찰했습니다.
서양 얼룩
단백질 발현을 평가하기 위해 TiO2에서 배양된 BMSC 나노튜브는 트립신(Gibco)으로 수확되었습니다. 세포를 PBS로 3회 세척하고 프로테아제 및 포스파타제 억제제 칵테일이 보충된 RIPA 완충액으로 얼음 위에서 30분 동안 용해시켰다. 용해물을 12,000xg에서 원심분리하여 수집했습니다. 4°C에서 15분 동안 상등액의 총 단백질 농도는 BCA(bicinchoninic acid) protein assay kit(Beyotime)을 사용하여 제조사의 지시에 따라 측정하였다. 로딩 버퍼를 위와 같이 단백질 샘플에 첨가한 다음, 95°C에서 15분 동안 끓였습니다. Western blotting 분석을 위해 10μL의 단백질 제제를 12.5% SDS-PAGE 젤(EpiZyme Inc., Cambridge, MA, USA)에 로딩하고 120V에서 1시간 동안 전기영동한 다음 폴리비닐리덴디플루오라이드에 전기이동했습니다. (PVDF) 멤브레인을 250mA에서 2시간 동안. 그런 다음 멤브레인을 실온에서 진탕기에서 1시간 동안 TBST에서 5-10% 무지방 건조 우유로 차단하고 4°C에서 밤새 희석 완충액(Beyotime)에 희석한 1차 항체와 함께 인큐베이션했습니다. 다음으로, TBST로 5분 동안 3회 세척한 후 희석 완충액에 희석한 형광 결합 이차 항체를 첨가한 후 암실에서 1시간 동안 실온에서 배양하였다. 단백질 밴드는 2색 적외선 형광 이미징 시스템(Odyssey, LiCor Biosciences, Lincoln, NE, USA)에 의해 검출되었습니다. 특히, 내부 참조 단백질의 밴드가 통일된 경우 멤브레인을 벗겨내고 다른 1차 항체로 재프로브한 후 동일한 과정을 수행하였다. 우리는 GAPDH 항체를 내부 참조 단백질로 사용하여 단백질 발현을 정상화했으며 이 연구에 사용된 다른 1차 항체는 항-빈쿨린(1:1000 희석, Abcam, Cambridge, MA, USA), 항-FAK(1:1000 희석)였습니다. , Cell Signaling Technology, Danvers, MA, USA), 항-Runx2(1:1000 희석, Cell Signaling Technology), 항-RhoA(1:1000 희석, 세포 신호 기술), 항-F-액틴(1:500 희석) , Abcam), anti-Osx(1:500 희석, Abcam) 및 anti-pYAP(1:1000 희석, Cell Signaling Technology). 이차 항체는 염소 항-마우스 IgG H&L(IRDye® 680RD, 1:5000 희석, Abcam) 및 염소 항-토끼 IgG H&L(IRDye® 680RD, 1:5000 희석, Abcam)이었습니다.
정량적 실시간 PCR
7일째에 정량적 실시간 PCR을 수행하여 runt-related transcription factor 2(Runx2), Osterix(Osx), Alp, osteocalcin(OCN), RhoA, YAP, TAZ, vinculin(VCL), TiO2의 골형성 배지에서 성장한 세포의 초점 접착 키나제(FAK) 및 거핵모세포 백혈병 1(MKL1) 나노튜브. Total RNA Kit(R6812-01HP, Omega Bio-Tek Inc., Norcross, GA, USA)를 사용하여 세포에서 총 RNA를 추출했습니다. RNA 시료의 농도와 순도는 260파장에서의 광학밀도로 결정하였으며, A260/280 비율과 A260/230 비율이 모두 1.8 이상인 시료만을 분석하였다. RNA 샘플은 제조업체의 지침에 따라 qScript cDNA Synthesis kit(Takara, Shiga, Japan)를 사용하여 cDNA로 역전사되었습니다. 정량적 실시간 PCR은 QuantStudio 6 Flex 실시간 PCR 시스템(Life Technologies)을 사용하여 SYBR® Premix Ex Taq™(Takara)로 수행되었습니다. 하우스키핑 유전자인 GAPDH를 내부 참조로 사용하였다. 데이터는 비교 Ct(2
-ΔΔCt
) 방법으로 대조군과 비교하여 배수 변화로 표현하였다. 사용된 프라이머의 서열은 표 1에 나열되어 있습니다.
통계 분석
모든 데이터는 달리 표시되지 않는 한 3중 샘플을 사용한 최소 3회의 독립적인 실험을 나타냅니다. 데이터는 평균 ± 표준 편차(SD)로 표시됩니다. 그룹 간의 차이는 Student-Newman-Keuls 사후 테스트 또는 Student의 t에 이어 일원 분산 분석으로 평가되었습니다. 테스트. 피 값 <0.05는 통계적으로 유의한 것으로 간주되었습니다.
<섹션 데이터-제목="결과">
결과
표면 특성화
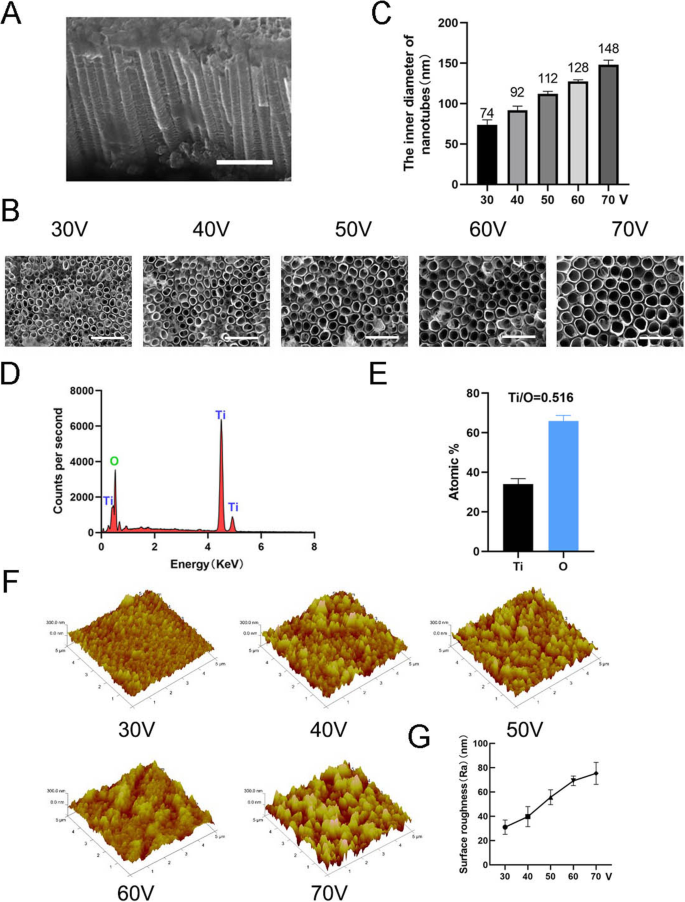
나노토포그래피를 제작하기 위해 TiO2 나노튜브는 1시간 동안 서로 다른 정전압(30, 40, 50, 60, 70V)에서 양극 산화 장비를 사용하여 순수한 티타늄 기판에 형성되었습니다(그림 1). 자기조립된 나노튜브의 균일하게 분포된 어레이는 주사전자현미경(SEM)에 의해 관찰되었다. 나노튜브의 측면 및 평면도는 그림 2a, b에 나와 있습니다. 이 연구의 모든 샘플에서 나노튜브의 높이는 약 2 μm인 반면, 나노튜브의 내부 직경은 약 74 nm(30 V), 92 nm(40 V), 112 nm(50 V), 128 nm(60 V) 및 148nm(70V)(그림 2c). 이는 나노튜브의 높이가 양극 산화 시간과 관련이 있고 내경이 양극 산화 전압과 관련이 있음을 나타냅니다. 그런 다음 X선 에너지 분산 분석(EDS)을 수행하여 나노튜브의 원소 조성을 분석했습니다. 이것은 나노튜브가 O와 Ti의 두 가지 요소로만 구성되어 있음을 보여주었습니다(그림 2d, e). AFM(Atomic Force Microscopy)을 사용하여 나노 튜브 구조를 감지하고 나노 튜브 프로파일의 산술 평균 편차를 표면 거칠기(Ra)로 측정했습니다(그림 2f). 데이터는 나노튜브의 표면 거칠기가 직경(즉, 양극 산화 전압)이 증가함에 따라 증가함을 보여주었다(그림 2g).
<그림>
나노튜브의 표면 특성화. 아 나노튜브의 측면도. 스케일 바:1μm. ㄴ 나노튜브의 평면도. 스케일 바:500 nm. ㄷ 5가지 다른 정전압(30, 40, 50, 60, 70V)에서 나노튜브의 내경. d 나노튜브 구조의 EDS 화학 원소 조성. 이 Ti와 O의 원소비. f 나노튜브 구조의 원자력 현미경(AFM) 이미지. 지 나노튜브의 평균 표면 거칠기(Ra)
TiO2 나노튜브로 유도된 MSC의 골형성 분화
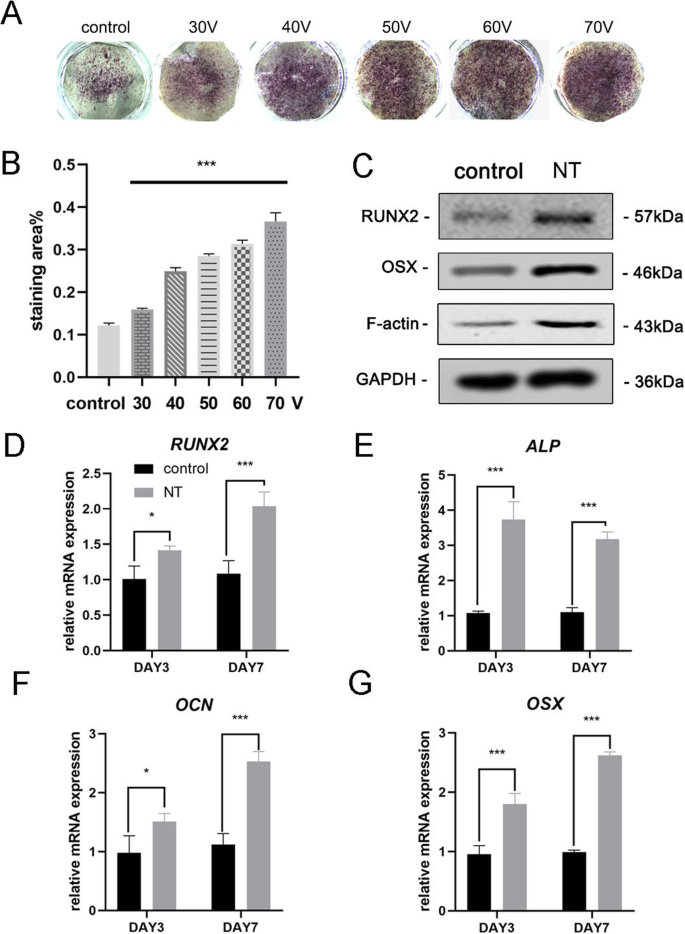
골유도 7일 후, MSC의 골형성 분화를 평가하기 위해 먼저 ALP 염색을 수행하였다. 염색 결과는 MSC가 TiO2에서 배양되었음을 보여주었습니다. 나노튜브는 매끄러운 티타늄 기질(대조군)에서 배양된 세포보다 더 높은 ALP 활성을 가졌다(그림 3a). 염색 영역의 통계적 분석은 대조군과 비교했을 때 골 형성 분화를 유도하는 나노튜브의 능력이 상당히 향상되었음을 보여주었다. 한편, 우리는 이 실험의 직경 범위 내에서 TiO2의 직경이 더 크게 되는 경향을 관찰했습니다. 나노튜브는 골형성 분화를 유도하는 능력이 더 강합니다(그림 3b). 따라서 70V 그룹은 결과를 더 잘 표시하기 위해 후속 실험에서 사용되었습니다. 다음으로, 우리는 3일과 7일에 골형성 유전자 발현을 분석했습니다. TiO2에서 배양된 MSC 3일 및 7일 동안 나노튜브는 대조군과 비교하여 골형성 유전자(RUNX2, ALP, OCN 및 OSX) 발현의 유의한 촉진을 보여주었다(그림 3d-g). Western blot 결과는 골유도 7일 후에 RUNX2와 OSX의 단백질 발현도 증가함을 확인하였다(Fig. 3c). 흥미롭게도 우리는 F-액틴이 TiO2에서 상향조절된다는 것을 발견했습니다. 나노튜브 그룹. 따라서 TiO2 나노튜브는 나노튜브의 직경과 관련된 조골세포 분화로 MSC를 지시했습니다. 우리의 결과는 또한 이 과정에서 F-액틴의 관여를 시사했습니다.
<그림>
TiO2 나노튜브는 BMSC의 골형성 유전자 발현을 향상시켰습니다. 아 매끄러운 티타늄 기질과 5가지 다른 나노튜브 기질의 ALP 염색. 세포를 7일 동안 골형성 배지로 유도하였다. ㄴ ImageJ를 사용하여 염색 영역의 통계 분석을 수행했습니다. ㄷ MSC의 골형성 관련 단백질(RUNX2 및 OSX) 및 F-액틴은 7일째에 웨스턴 블롯팅으로 분석되었습니다. RUNX2의 mRNA 발현(d ), ALP(e ), OCN(f ) 및 OSX(g ) 3일 및 7일에 qRT-PCR로 분석했습니다. NT 나노튜브 그룹. 데이터는 3개 샘플의 평균 ± SD를 나타냅니다. *피 <0.05, **P <0.01 및 ***P <0.001
TiO에서 MSC의 F-액틴 매개 조골세포 분화2 나노튜브
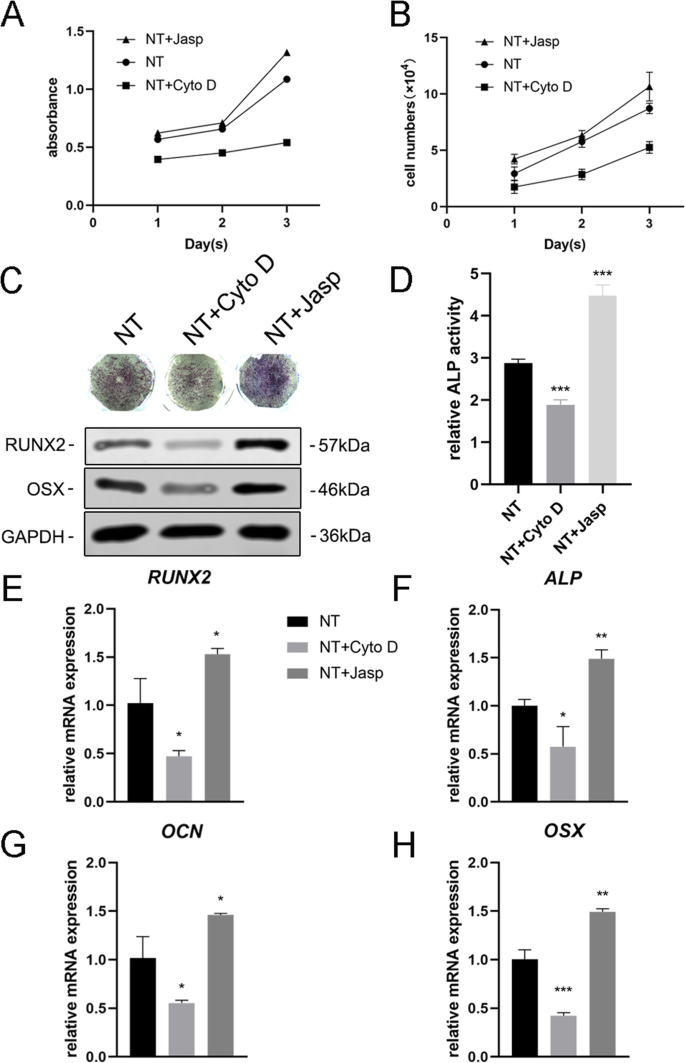
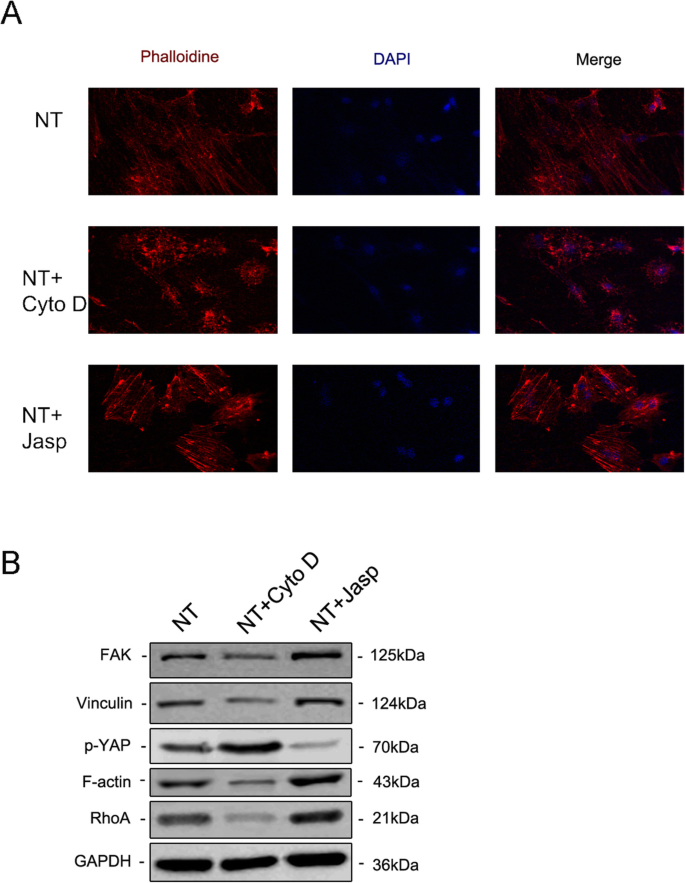
F-액틴이 나노토포그래피로 유도된 세포 분화에 관여하는지 여부를 추가로 조사하기 위해 jasplakinolide(Jasp) 및 cytochalasin D(Cyto D)의 두 시약을 사용하여 F-액틴 중합을 각각 양성 및 음성 방식으로 조절했습니다. Rhodamine-Phalloidin 염색의 공초점 현미경 사진은 Cyto D 처리군의 F-actin이 거의 해중합되어 섬유질 구조가 거의 보이지 않는 반면 Jasp가 안정화되고 중합된 F-actin은 관찰된 것보다 더 뚜렷하고 밝은 번들 같은 구조로 확인됨을 보여주었습니다. 대조군에서(그림 5a). 또한 Western blot 분석에서도 F-actin의 단백질 발현이 영향을 받는 것으로 확인되어 Cyto D와 Jasp가 모두 예상대로 작용함을 보여주었다(Fig. 5b). 세포 증식 분석은 Cyto D가 세포 증식을 유의하게 억제한 반면 Jasp는 세포 성장을 촉진함을 보여주었습니다(그림 4a). 세포 수 결과는 이 발견과 일치했습니다(그림 4b).
<그림>
F-액틴 어셈블리는 BMSC에서 골형성 유전자의 발현을 조절했습니다. 아 , b Cyto D 및 Jasp 처리 후 세포 증식은 1일, 2일 및 3일에 CCK-8 분석 또는 자동화된 세포 계수기를 사용하여 결정되었습니다. c , d 3일 동안 약물 처리 후 MSC에서 ALP의 발현을 평가하기 위해 ALP 염색 및 ALP 활성 분석을 수행하였다. ImageJ를 사용하여 염색 영역을 분석했습니다. ㄷ , e –h NT+ Cyto D 그룹과 NT+ Jasp 그룹의 골형성 관련 마커의 변화를 대조군(약물 처리 없음)과 비교하기 위해 Western blotting과 qRT-PCR을 사용했습니다. NT 나노튜브 그룹. 데이터는 3개 샘플의 평균 ± SD를 나타냅니다. *피 <0.05, **P <0.01 및 ***P <0.001
다음으로, 우리는 F-액틴이 이 과정을 매개하는지 여부를 조사하기 위해 MSC가 조골세포로 분화하는 능력을 평가했습니다. 우리는 먼저 ALP를 골형성의 초기 마커로 검출했습니다. 대조군과 비교하여 Cyto D 처리는 ALP의 발현과 그 활성을 감소시켰고 Jasp 처리군은 상향 조절되었다(그림 4c, d). 이 결과와 일치하게, Jasp 처리는 RUNX2 및 OSX의 단백질 수준을 증가시키는 반면 Cyto D는 반대 효과를 나타냈습니다(그림 4c). 이와 일치하여 RUNX2, ALP, OCN 및 OSX를 포함한 골 특이적 유전자의 mRNA 발현 수준은 약물 치료 후 동일한 경향을 보였다(그림 4e-h). 무엇보다 이러한 데이터는 F-actin이 TiO2에 의해 유도된 MSC의 골형성 분화 과정에서 중요한 역할을 함을 나타냅니다. 나노튜브. F-액틴 해중합을 촉진하면 나노토포그래피에 의한 조골세포 분화가 억제되는 반면, F-액틴의 안정화 및 중합은 조골세포 분화를 강화합니다.
TiO에서 MSC의 F-액틴 조절 조골 세포 분화2 MKL1 및 YAP/TAZ를 통한 나노튜브
MSC의 운명 조절에 F-액틴과 관련된 기본 메커니즘을 해부하기 위해 우리는 F-액틴과 직접 상호작용하거나 F-액틴 중합에 영향을 미치는 단백질/분자를 조사했습니다. 먼저, 우리는 나노토포그래피가 F-액틴과 G-액틴 사이의 균형에 어떻게 영향을 미치는지 확인하려고 했습니다. TiO2 물리적 신호로서의 나노튜브는 막 투과성 화학 신호와 다르므로 자극을 세포로 전달하기 위해 세포막의 일부 구성요소를 사용해야 합니다. 축적된 증거는 인테그린, 탈린, 초점 접착 키나제(FAK), 빈쿨린(VCL), 텐신 및 기타 단백질을 포함하는 국소 접착 복합체가 세포외 기질(ECM)의 상태에 대해 세포에 알리는 신호 전달자 역할을 함을 나타냅니다. ) 따라서 생물학적 행동에 영향을 미칩니다[22, 23]. 더 중요한 것은 F-액틴이 이러한 국소 접착 복합체를 통해 인테그린에 결합하여 세포내 액틴 번들과 ECM 사이에 기계적 연결을 형성한다는 것입니다[24]. 결과적으로, 우리는 다음으로 초점 접착 복합체의 구성 요소 표현을 분석했습니다. 결과는 VCL 및 FAK의 단백질 및 mRNA 발현이 F-액틴의 변화와 일치함을 보여주었으며, 이는 초점 접착 복합체가 TiO2에 의해 유도된 MSC의 골 형성 분화 과정에 관여했음을 나타냅니다. 나노튜브(그림 5b 및 6a, b). 또한, 우리는 GTPase의 Rho 계열에 있는 작은 GTPase 단백질인 RhoA가 Jasp 치료 그룹에서 상향 조절되고 Cyto D에 의해 억제된다는 것을 발견했습니다(그림 5b 및 6a, b). RhoA는 MAPK 경로에서 중요한 상류 신호 전달 분자이며 FAK에 의해 조절될 수 있습니다[25, 26]. RhoA의 주요 기능은 응력 섬유(F-액틴)의 중합 및 안정성과 초점 접착 복합체의 조립을 촉진하는 것입니다[27]. 종합하면 이 데이터는 TiO2 나노튜브는 초점 접착 복합체와 RhoA를 통해 F-액틴 중합에 영향을 미칠 수 있습니다.
<그림>
면역형광 염색은 로다민 결합 팔로이딘(a ). The protein expression of FAK and VCL contained in the focal adhesion complex, RhoA and phosphorylated YAP were investigated by Western blotting (b ). NT the nanotubes group
Effect of F-actin assembly on FAK (a ), vinculin (VCL) (b ), RhoA (c ), MKL1 (d ), YAP (e ), and TAZ (f ) gene expression in MSCs. NT the nanotubes group. Data represent the mean ± SD of three samples. *피 <0.05, **P <0.01, and ***P <0.001
But how does F-actin regulate cell fate? Most studies have demonstrated that F-actin is involved in cell migration, cell division, endocytosis, and especially tumor cell invasion [28,29,30]. Few studies have suggested that F-actin could also regulate cell differentiation, let alone its specific molecular mechanism [31, 32]. Consequently, we searched for articles that mentioned the F-actin changes and found that YAP/TAZ, two closely related transcriptional co-activators in the Hippo signaling pathway, which shuttle between the cytoplasm and the nucleus, may serve as mechanotransducers in regulating MSC differentiation [33,34,35]. In addition, we also found that MKL1, a key regulator of smooth muscle cell differentiation, which interacts with the transcription factor serum response factor, could bind to G-actin and also circulate between the cytoplasm and the nucleus [21, 36]. Our results ultimately identified the involvement of YAP/TAZ and MKL1 in nanotube-induced osteoblast differentiation mediated by F-actin (Figs. 5b and 6d–f). Interestingly, the protein expression of phosphorylated YAP showed the opposite trend, indicating that not only was the expression of YAP changed, but the phosphorylation of YAP was also changed by Cyto D and Jasp (Fig. 5b). This result was consistent with the report that the phosphorylation of YAP/TAZ could be sequestrated in the cytoplasm [35].
In summary, our results preliminarily demonstrated that F-actin regulated osteoblast differentiation of MSCs on TiO2 nanotubes through MKL1 and YAP/TAZ (Fig. 7).
Schematic representation of F-actin assembly induced by nanotubes, and the putative role of MKL1 and YAP/TAZ in acting as the downstream mediators of F-actin signaling to regulate gene expression
토론
Titanium and titanium alloys are the most widely used metal materials in orthopedic clinical implants due to the good properties of titanium [3]. However, aseptic loosening is still an urgent problem to be solved and improved, and the key is likely to lie in improving the integration of the implant and the host bone. Previous studies have shown that surface coating and modification or immobilization of biofunctional molecules will be beneficial to osseointegration [37]. Recently, the surface topography of implants has attracted the attention of many researchers thanks to studies into the cell response to physical cues [9,10,11, 13, 38]. In this study, we demonstrated the ability of nanotubes to promote osteogenic differentiation of MSCs, and this ability was enhanced with increasing inner diameter of the nanotubes (30–70 V). This will help guide the diameter of the nanotubes on the surface of the implants.
As a topographical structure, nanotubes first change the physical properties of the material, such as adsorption capacity and electrical and thermal conductivity. These physical properties determine their application in the industrial field. For example, most high-voltage power transformers need to be filled with insulating material, which is usually transformer oil or insulating gas. When the insulation of a transformer fails due to overheating and partial discharge, a serious discharge accident will occur. Therefore, finding an effective method that accurately detects the concentration and types of dissolved gases or insulating gas decomposition components in a transformer is necessary to monitor the operating state of the transformer [8, 39,40,41]. The traditional approach is to look for materials with good gas adsorption in transition elements, which are rich in d electrons, such as Pd(1 1 1) [39]. Nowadays, nanotubes are widely studied for their good gas adsorption properties. 그는 외. found that CuO-BNNT was suitable for the adsorption of C2 H2 , because of its stronger adsorption on C2 H2 [8]. Meanwhile, TiO2 itself can be a gas-sensing material. Gui et al. found that Co-doped TiO2 further enhanced gas adsorption capacity and exhibited a superior adsorption ability and conductivity change toward C2 H4 molecules [40]. Consistent with this study, Mn-doped graphene also exhibited enhanced conductivity and superior capability of C2 H2 and CO detection than pristine graphene [41]. The above research indicates that the TiO2 nanotubes prepared in our experiment have a potential application in the field of monitoring the operative state of a transformer. However, the adsorption capacity and electrical conductivity of the nanotubes to gases need to be further studied, especially whether these properties are enhanced after doping with transition elements (e.g., Mn).
In addition to changing the physical properties of a surface, nanoscale morphology also affects the biological behavior of the cells attached to it. Cells first adhere to the surface of the material and then migrate, proliferate, and differentiate. Compared to a flat surface, the hollow structure of the nanotubes provides fewer adhesion sites for cells. Therefore, in order for the cells to adhere to the nanotube surface steadily and maintain the biomechanical balance within the cell, the focal adhesion complex begins to assemble and mature, and F-actin becomes strong and stable.
F-actin, a linear polymer microfilament consisting of G-actin monomers, is one of the three major components of the cytoskeleton. As a mechanical-loading structure, F-actin is generally believed to be involved in cell division, cell migration, endocytosis, and tumor cell invasion [28,29,30], but some recent studies showed that it can also affect cell differentiation [31,32,33, 36]. For example, actin cytoskeletal depolymerization by simvastatin induces chondrocyte differentiation [31], and actin depolymerization enhances adipogenic differentiation in human stromal stem cells [32]. Our results also revealed that, compared with the control group, MSCs cultured on nanotubes had higher F-actin levels and a more obvious fibrous structure. Meanwhile, promotion of F-actin polymerization by Jasp enhanced osteogenic differentiation, while the depolymerization of F-actin inhibited osteogenic differentiation, suggesting that F-actin mediates TiO2 nanotube-induced osteoblastic differentiation of MSCs.
F-actin can be regulated by Rho GTPases, members of the Ras superfamily [23, 42], and Rho can induce actin reorganization through at least two effectors, ROCK and Dia. ROCK is activated by binding to Rho-GTP and then myosin light chain (MLC), the substrate of ROCK, plays an important role in F-actin assembly. ROCK inhibits the activity of MLC phosphatase, leading to an increase in MLC phosphorylation, which stimulates the ATPase activity of myosin II and promotes the assembly of F-actin. In addition, ROCK also targets LIM kinase (LIMK). Phosphorylated LIMK inactivates cofilin by phosphorylation, which can disassemble F-actin in its active state. Another effector is Dia, a member of the formin-homology (FH) family of proteins which contains two FH domains. These domains contain multiple proline-rich motifs which bind to the G-actin-binding protein, profilin. This interaction contributes to actin polymerization and F-actin organization [42]. We detected one of the Rho GTPases, RhoA, and found that the expression of RhoA was consistent with the level of F-actin. However, we were unable to clearly describe how the nanotubes regulate the expression of RhoA, because there are many other regulators, including integrin signaling, other adhesion receptors, G protein-coupled receptors (GPCRs), soluble factors such as LPA, receptor tyrosine kinase signaling, and so on [43].
Knowing that F-actin can be regulated by RhoA, we next asked what role focal adhesion played in this process, because focal adhesion complexes, containing integrins, talin, vinculin, paxillin, and focal adhesion kinase (FAK), are formed and mature when cells attach to the surface of nanotubes. Integrins are transmembrane heterodimers that couple the ECM to the other focal adhesion proteins so as to facilitate cell attachment. They not only act simply as hooks but also transmit to the cell critical signals about the nature of its surroundings, which along with other signals such as EGFR, prompt the cell to make decisions about its biological behaviors. These signals are further transmitted to F-actin, which is directly connected to the focal adhesion complexes. On the one hand, the nanoscale morphology causes focal adhesion complex assembly and maturation. On the other hand, kinases such as FAK and Src kinase family members will recruit molecules such as CRK to self-regulate the assembly and maturation of focal adhesion complexes [44,45,46]. Our results demonstrated that the formation and maturation of focal adhesion complexes were impaired by F-actin depolymerization, suggesting that there was a feedback from focal adhesion complexes to actin assembly in line with published reports.
However, it should not be ignored that these proteins contained in focal adhesion complexes have the function of signal transduction [47]. That is to say, nanotubes may directly regulate gene expression through signal cascades, and F-actin may just participate in or be affected by this process. For instance, the dual kinase complex of FAK and Src can regulate Rho GTPases such as RhoA. This shows that nanotubes can regulate RhoA through integrins and the FAK/Src complex. In addition Src, a non-receptor tyrosine kinase protein, can activate Ras (small GTPase) by phosphorylating FAK at tyrosine residue 925 [47, 48]. Then, Ras activates numerous biochemical pathways, including the well-studied MAPK pathway and the PI3K/AKT/mTOR pathway. In the MAPK pathway, Ras activates c-Raf, followed by mitogen-activated protein kinase kinase (MAP2K) and then MAPK1/2, also known as extracellular signal-regulated kinase (ERK). ERK in turn activates transcription factors such as serum response factor (SRF) and c-Myc that are involved in regulating growth and differentiation [49]. What is more, Runx2, a key transcription factor in osteogenic differentiation, can also be regulated by ERK [50], and our previous study confirmed that mechanical strain promoted osteogenic differentiation of BMSCs through the FAK-Erk1/2-Runx2 pathway [17]. Therefore, we cannot rule out that ERK plays a role in nanotube-induced osteogenic differentiation and further study is still needed.
So what exactly is the role of F-actin in inducing differentiation of nanotubes, because its change can affect cell differentiation? One possibility is that the change of F-actin assembly can inversely regulate the level of FAK so as to induce osteogenic differentiation through the FAK-Erk1/2-Runx2 pathway as described above, because in our results, focal adhesion complexes and actin polymerization showed the same trends of change, indicating that they act as a whole in response to the extracellular environment. However, some other possibilities also exist, and a number of articles have shown that MKL1 and YAP/TAZ act downstream of the actin dynamic balance [20, 51,52,53,54]. Both of them shuttle between the cytoplasm and the nucleus, and may help to transduce signals from the cytoskeleton to the nucleus.
MKL1, also termed myocardin-related transcription factor A, is sensitive to changes in G-actin levels. When cytoplasmic G-actin levels increase, monomeric G-actin binds to MKL1 and prevents it from binding to SRF and activating transcription. SRF target genes include actins such as smooth muscle actin (SMA) as well as other actin-binding proteins, including immediate early genes like c-fos and egr1. Recent studies have demonstrated that changing SRF activity could regulate adipogenesis by activating the adipogenesis transcription factor peroxisome proliferator-activated receptor γ (PPARγ), and also regulate bone formation via IGF-1 and Runx2 signaling [55, 56].
YAP and TAZ are two transcriptional coactivators in the Hippo signaling pathway, identified as an important regulatory pathway that restricts cell proliferation, thereby controlling organ size and morphogenesis [20]. Large tumor suppressor genes 1 and 2 (LATS1/2) phosphorylate them, thereby creating a binding site for 14-3-3 proteins, the binding of which prevents their nuclear import [53, 54]. As a consequence, phosphorylated forms of YAP/TAZ are sequestered in the cytoplasm, preventing the expression of genes like Ctgf and Areg. In addition, some studies have shown that YAP/TAZ can interact with T-box 5 (TBX5), RUNX2, and p73 to regulate gene expression [57,58,59]. Further, cell adhesion to cell matrix proteins has been shown to trigger YAP nuclear localization through an integrin/FAK/Src axis. In our study, the results suggested that this pathway was possibly involved in nanotube-induced differentiation. Further study into the downstream mediators of the integrin/FAK/Src axis should be carried out to clarify the specific mechanism.
On the other hand, more and more studies illustrate that F-actin interacts with Hippo signaling, and somehow inhibits the phosphorylation of YAP [54, 60], which is consistent with our experimental results that promoting F-actin polymerization reduces the expression of phosphorylated YAP. We hypothesize that ATP involved in the process of the transformation between G-actin and F-actin may also play an important role in the phosphorylation of YAP, which is yet to be studied.
After understanding the above possible molecular mechanisms, we can try to explain some of the experimental phenomena found in this study. Our results revealed that the larger the diameter of the nanotubes, the stronger the ability of the nanotubes to promote osteogenic differentiation. This is consistent with previous research [61, 62]. The reason for this phenomenon is that the larger the diameter of the nanotubes, the less adhesion sites they can provide to the cells, and the greater the assembly and maturity of focal adhesion complexes. Along with these, stress fibers made of F-actin will have greater strength and stability. These structures enhance the signaling that promotes osteogenic differentiation. Predictably, however, this effect is significantly reduced when the nanotubes become too large in diameter, making it difficult for the cells to adhere to the surface [12]. Similarly, when the height of the nanotubes is inconsistent, the differences in height can result in a change of adhesion site and rearrangement of the cytoskeleton, which will further affect cell differentiation. Intriguingly, even flat surface materials without nanotube modification can induce changes in cell differentiation. A number of studies have demonstrated that focal adhesion formation and stress fiber organization are regulated by substrate stiffness [63,64,65], and YAP/TAZ also plays an important role in this process. Therefore, it is obvious that the integrins–FAs (focal adhesions)–F-actin axis plays a role in the transduction of physical signals into intracellular chemical signals.
In summary, our results demonstrated that F-actin regulates osteoblastic differentiation of mesenchymal stem cells on TiO2 nanotubes through MKL1 and YAP/TAZ, whose target genes partly explained the proliferation and differentiation of MSCs. We know that there is no single change in the signal network and any change is regulated by numerous molecules and proteins. One type of biological behavior must be the result of the regulation of a series of signaling pathways. Nanotubes induce cell differentiation by triggering a complex network of signals, including integrins, proteins contained in focal adhesion complexes, FAK, Src, Rho GTPase, the MAPK pathway, the Hippo pathway, and other reported signaling pathways. At least as important, there are many signal cycles in the signal network and a downstream signal can regulate the upstream signal via feedback. In this study, we found that vinculin and FAK can be regulated backwards by F-actin assembly, increasing the uncertainty of molecular function. Therefore, more details of the molecular mechanism await further study.
결론
Our results showed that TiO2 nanotubes promoted the osteogenic differentiation of MSCs, and this ability was enhanced with the increasing diameter of nanotubes within a certain range (30–70 V). F-actin mediated nanotube-induced cell differentiation through MKL1 and YAP/TAZ, providing a novel insight into the study of cell differentiation.
데이터 및 자료의 가용성
현재 연구 중에 사용 및 분석된 데이터 세트는 합당한 요청이 있는 경우 교신저자에게 제공됩니다.