

나노물질

산업 제조
나노하이드록시아파타이트(nano-HA)는 재생의학 분야에서 상당한 주목을 받고 있다. 내피 세포(EC)-중간엽 줄기 세포(MSC) 상호 작용은 뼈 재건에 필요하지만 나노 HA가 이 과정에서 상호 작용하는 방식은 아직 알려지지 않았습니다. 여기에서 우리는 EC에 의해 중재되는 간접 공동 배양 모델을 사용하여 MSC에 대한 HA 나노 입자(HANP)의 세포 독성 및 골유도 효과를 조사하고 기본 메커니즘을 강조했습니다. 준세포독성 용량에서 HANP는 MSC의 광물화된 결절 및 알칼리성 포스파타제 생산뿐만 아니라 조골 세포 유전자의 생존력 및 발현을 증가시키는 것으로 밝혀졌습니다. 이러한 현상은 EC에서 분비되는 HIF-1α에 의존하여 ERK1/2 신호 전달 캐스케이드를 촉발했습니다. 또한 HIF-1α가 MSC의 골 형성 분화에 미치는 영향을 정량적으로 분석하기 위해 2단계 세포 계통 수학적 모델을 수립했습니다. HIF-1α가 최대 1500pg/mL까지 MSC의 골형성 분화율에 대해 용량 의존적 자극 효과를 발휘하는 것으로 나타났으며 이는 위의 결과와 일치했습니다. 우리의 데이터는 HNP, EC 및 MSC 간의 협력 상호 작용이 뼈 재생을 자극하는 역할을 할 수 있음을 시사했습니다. 또한, 2단계 세포 계통 모델은 뼈 조직 공학에서 이펙터 분자의 잠재적 영향을 평가하기 위한 시험관 내 시스템에 유용합니다.
외상, 선천적 기형 또는 외과적 절제로 인한 골 결손의 재건은 정형외과 수술에 있어 큰 도전 과제이다[1]. 대표적인 생리활성 세라믹인 수산화인회석(Hydroxyapatite, HA)이 골대체재로 사용되어 왔다[2]. 그러나 바람직하지 않은 기계적 및 골유도 특성으로 인해 임상 적용이 제한됩니다[3]. 최근 몇 년 동안 nano-HA는 고유한 생체 학적 특성으로 인해 최적의 더 나은 생체 활성과 향상된 기계적 성능을 보여 주며 재생 의학과 관련된 생물 의학 분야에서 상당한 관심을 받고 있습니다 [4]. nano-HA를 뼈 결함에 이식하면 뼈 복구에 관여하는 여러 세포가 노출됩니다. 따라서 nano-HA의 생물학적 거동을 평가할 필요가 있습니다. 여러 증거에서 HA 나노입자(HANP)가 인간 탯줄 Wharton의 젤리 유래 중간엽 줄기 세포(hWJ-MSC) 및 조골 세포에 의해 흡수되어 골 형성 분화가 향상될 수 있음을 직접 입증했습니다[5,6,7]. Duaet al. 이전에 공학된 연골을 새로운 연골로 통합하는 것을 촉진하는 HANP의 능력이 보고되었습니다[8]; 반대로, HAP는 인간 제대 정맥 내피 세포(HUVEC)의 혈관 신생 능력을 억제합니다[9]. 인간 건강 측면에서 HNP가 뼈 재생에 미치는 영향에 대한 보다 포괄적인 이해가 필요하며, 공학 나노 기반 인공 뼈의 지속적인 적용은 이러한 연구의 시급성을 더합니다.
뼈 재생은 필연적으로 신생혈관의 침범을 동반합니다. EC는 수동적으로 혈액을 전달하는 역할을 하고 기관 재생을 유도, 지정 및 안내하고 항상성과 신진대사를 유지하는 역할을 하는 혈관계의 내부 세포 라이닝입니다[10, 11]. MSC는 내피 주위 틈새의 일부이며 상주하는 틈새 내에서 특정 생리학적 및 생화학적 미세 환경의 유도 하에 자가 재생 및 다중 분화 능력을 보유합니다[12]. Tsai et al. EC가 엔도텔린-1을 분비하여 MSC가 골 및 연골 계통 분화를 유도할 수 있음을 발견했습니다[13]. 또한 Saleh et al. HUVEC 분비 단백질 및 MSC 막 결합 수용체와 상호작용하여 증식 및 골형성 분화를 향상시키는 관련 누화 신호 전달 경로를 식별하기 위해 마이크로어레이 데이터 분석을 사용했습니다[14]. 뼈 조직 공학에서 HAP는 신생혈관과 접촉할 수 있고 EC에 의해 세포내이입될 수 있으며, 이는 이러한 세포의 생리학적 기능을 변경하는 것으로 나타났습니다[9, 15]. 이것은 또한 주변 골전구 세포에 영향을 미치고 측분비 신호를 변경하여 뼈 복구에 영향을 줄 수 있습니다. 그러나 HANP가 MSC에 미치는 직접적인 영향이 조사되었지만 HANP가 EC를 통해 MSC의 골형성 분화를 간접적으로 유도할 수 있는지 여부에 대한 명확한 이해가 여전히 부족합니다. 뼈 수선에.
이 연구에서는 EC와 MSC 간의 상호 작용에 대한 HAP의 생물학적 효과에 대한 추가 통찰력을 얻기 위해 HUVEC 및 hWJ-MSC를 사용하여 간접 공동 배양 모델을 설정했습니다. 이 시스템을 활용하여 HUVEC 매개 측분비 신호를 통한 hWJ-MSC에 대한 HAP의 세포 독성 및 골유도 효과를 평가했습니다. HANP-유도 내피 세포-MSC 상호 작용에 영향을 미치는 주요 요인을 확인하기 위해 HANP로 자극된 HUVEC의 상층액에 있는 가용성 인자를 유전자 및 단백질 수준에서 관련 메커니즘에 중점을 두고 평가했습니다. 결과는 저산소증 유발 인자(HIF)-1α가 이러한 상호 작용에서 중요한 역할을 한다는 것을 보여주었습니다.
HIF-1α가 골형성 과정에 미치는 영향을 정량적으로 관찰하고 예측하기 위해 2단계 세포 계통과 HIF-1α를 결합한 수학적 모델을 수립했습니다. 여기서, 경험적 데이터를 분석하여 정의된 초기 세포 파종 밀도 및 HIF-1α 농도를 기반으로 하여 임의의 시점에서 MSC 수 및 분화 정도를 예측하기 위해 2단계 세포 계통 모델을 사용하였고, 이는 차례로 초기 배양 조건 및 배양 시간에 대한 적절한 제안을 제공합니다. 이 연구의 결과는 재생 의학에 사용하기 위한 혁신적인 생체 재료의 개발을 촉진하는 역할을 할 수 있는 나노 기반 뼈 대체물과 생물학적 시스템 간의 상호 작용을 밝히는 데 도움이 될 것입니다.
20nm(np20), 20*80nm(np80)의 HANP 및 순도가 ≥ 99.0%인 초소형 HA 입자(m-HAP)는 Nanjing Emperor Nano Material Company Ltd(중국 난징)에서 구입했습니다. 입자의 크기와 모양은 투과전자현미경(TEM; FEI Tecnai G2 Spirit Bio-Twin, FEI, Hillsboro, OR, USA)과 주사전자현미경(SEM; LEO1530VP, Germany)을 사용하여 관찰하였다. HA 입자(HAP)의 유체역학적 크기와 제타 전위는 Zetasizer Nano ZS90 및 Mastersizer 3000(Malvern Instruments, Malvern, UK)을 통해 결정되었습니다.
모든 실험 프로토콜은 난징 의과 대학 윤리 위원회의 승인을 받았습니다. HUVEC 및 hWJ-MSC는 이전에 설명된 바와 같이 기증자의 서면 동의를 얻은 후 신선한 인간 탯줄에서 수확되었습니다[16, 17]. 간단히 말해서, 탯줄과 탯줄을 1% 페니실린과 스트렙토마이신(PS; Hyclone, GE Healthcare Life Sciences, Pasching, Austria)을 포함하는 인산완충식염수(PBS)로 헹구었습니다. 그런 다음 제대 정맥에 0.1% 콜라게나제 I(Sigma, St. Louis, MO, USA)을 채우고 37°C에서 15분 동안 배양했습니다. 수집 후 HUVEC를 EC 배지(ECM)(Sciencell, San Diego, CA, USA)에서 배양했습니다.
그 후 혈관을 제거하고 Wharton's jelly를 1mm 2 크기로 자릅니다. 조각으로 만든 다음 25cm 2 에 놓습니다. 조직 배양 플라스크(Corning Incorporated, Corning, NY, USA). 이 세포는 10% 소태아혈청(GIBCO) 및 1% PS가 보충된 L-DMEM(GIBCO Life Technology, Grand Island, NY, USA)에서 배양되었습니다.
hWJ-MSC는 CD13, CD29, CD34, CD44, CD45, CD51 및 CD105에 대한 단일클론 항체를 사용하여 표현형을 확인하기 위해 평가되었습니다(BD Biosciences, San Jose, CA, USA). HUVEC는 von Willebrand factor(vWF; Shanghai ChangDao Biotech Co, Ltd., Shanghai China)를 사용하여 평가되었습니다. 이 실험에서는 3-7절 사이의 HUVEC와 3-5절 사이의 hWJ-MSC를 사용했습니다.
그런 다음 PBS 중 1mg/mL의 입자 현탁액을 L-DMEM에서 최종 농도로 희석했습니다. 그림 1에서 볼 수 있듯이 HUVEC는 표시된 농도의 HAP에서 18시간 동안 배양되었습니다. 배양 배지를 4°C에서 15분 동안 15,000rpm으로 원심분리하고, 10% FBS가 보충된 상등액을 hWJ-MSC의 조건 배지(CM)로 사용하여 다음 실험을 수행했습니다. CM은 10mM β-글리세로포스페이트, 50μg/mL L-아스코르브산-2-포스페이트 및 0.1μM 덱사메타손(Sigma-Aldrich, St Louis, MO, USA)을 포함하는 골형성 유도액으로 보충된 골형성 배지로 구성되었습니다. 또한 2-methoxyestradiol(2-MeOE2)(Selleck Chemicals, Houston, TX, USA)을 특정 HIF-1α 억제제로 사용했습니다. 2-MeOE2(+) 그룹에서 hWJ-MSC는 HAP 보충 전에 40분 동안 5μM 2-MeOE2로 전처리된 HUVEC의 CM으로 배양되었습니다. PD98059는 특정 MEK 억제제로 사용되었습니다. PD98059 그룹에서 hWJ-MSC는 5μM PD98059를 포함하는 CM과 함께 배양되었습니다.
<그림>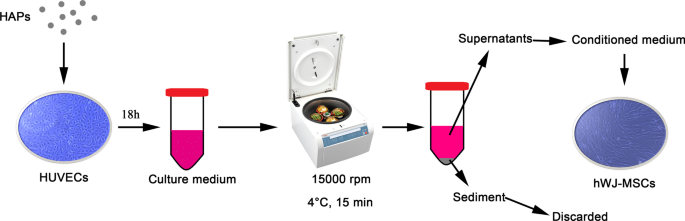
HUVEC에 의해 매개되는 HAP/hWJ-MSC 간접 공동 배양의 그림. 약어:HAP 수산화인회석 입자, hWJ-MSC 인간 탯줄 Wharton의 젤리 유래 중간엽 줄기 세포, HUVEC 인간 제대 정맥 내피 세포
MTS Assay Kit(MTS; Bestbio, Beyotime Biotechnology, Shanghai, China)를 사용하여 세포 생존율을 평가했습니다. hWJ-MSC를 24시간 동안 부착시킨 다음 CM과 함께 24시간 및 72시간 동안 배양했습니다. 포르마잔의 흡광도는 마이크로플레이트 판독기(SpectraMax M2; Molecular Devices LLC, Sunnyvale, CA, USA)를 사용하여 490nm에서 평가되었습니다. 흡광도도 같은 조건에서 표준 보정 곡선을 사용하여 세포 수로 변환되었습니다.
TRIzol 시약(Invitrogen, Carlsbad, CA, USA)을 사용하여 지정된 시간 동안 CM에서 배양된 hWJ-MSCs 세포에서 전체 RNA를 분리했습니다. T3 열순환기(Mastercycler 5333; Eppendorf, Hamburg, Germany)에서 PrimeScript First Strand cDNA Synthesis kit(TaKaRa, Tokyo, Japan)를 사용하여 1.0μg RNA에서 상보적 DNA를 전사했습니다. 표시된 유전자의 발현 수준은 정량적 실시간 증폭 시스템(7900HT Fast; Applied Biosystems, Foster City, CA, USA)에서 FastStart Universal SYBR Green Master(ROX) 키트(Roche, Basel, Switzerland)를 사용하여 분석되었습니다. 표적 유전자의 상대 mRNA 발현은 GAPDH로 정규화한 다음 \(2^{{ - \Delta \Delta C_{t} }}\) 방법을 사용하여 결정되었습니다. 표적 유전자에 대한 프라이머 서열은 표 1에 나열되어 있습니다.
hWJ-MSC를 12-well plate에서 osteogenic medium으로 최대 14일 동안 배양한 후 ARS 염색법(Leagene, Leagene Biotechnology, Beijing, China)을 이용하여 세포외 기질 광물화를 관찰하였다. 간단히 말해서, 샘플을 15분 동안 무수 에틸 알코올로 고정한 다음 실온에서 5분 동안 1%(w/v) ARS(pH, 4.2)로 염색했습니다. 염색된 세포를 이중 증류수로 2회 세척한 후 사진을 촬영하였다. 광물화 과정의 정량적 분석을 위해 300μL의 10%(w/v) 세틸피리디늄 클로라이드 일수화물(BOMEI, BOMEI Biotechnology, Hefei, China)을 각 웰에 첨가하고 플레이트를 30분 동안 인큐베이션했습니다. 각 샘플의 총 90μL를 96웰 플레이트로 옮기고 405nm에서 흡광도를 3회 측정했습니다.
hWJ-MSC를 골형성 배지가 있는 12웰 플레이트에서 최대 14일 동안 배양한 후 BCIP/NBT Alkaline Phosphatase Color Development Kit(Beyotime)를 사용하여 ALP 염색을 수행했습니다. 간단히 말해서, hWJ-MSC는 4% 파라포름알데히드에 고정되었습니다. 그런 다음 샘플을 nitro-blue tetrazolium과 5-bromo-4-chloro-3-indolyl-phosphate의 혼합물에서 4시간 동안 염색하고 사진을 촬영했습니다. ALP 합성을 정량화하기 위해 세포를 얼음 RIPA 용해 완충액(Beyotime)에서 30분 동안 용해했습니다. 세포 용해물을 4°C에서 10분 동안 12,000rpm으로 원심분리하고 상층액을 ALP 분석 키트(Beyotime)를 사용하여 ALP 정량 분석을 거쳤습니다. 흡광도는 405nm에서 3회 측정되었으며 표준 곡선을 사용하여 ALP 활성으로 변환되었습니다.
HUVEC는 2 × 10 5 에서 시딩되었습니다. 세포/웰. CM은 HAP와 함께 18시간 동안 배양된 HUVEC에서 수집하고(추가 파일 1), 상층액을 ELISA 분석했습니다(그림 1). 인간 HIF-1α ELISA 키트(Anhui Joyee Biotechnics, Anhui, China)는 제조업체의 지침에 따라 사용되었습니다.
HUVEC를 12웰 플레이트의 슬라이드에 접종했습니다(7.6 × 10 4 세포/웰). CM에 18시간 노출된 후 세포를 4% 파라포름알데히드(Biosharp, Beijing, China)에 고정하고 PBS 중 0.1% Triton X-100(Beyotime)으로 투과화한 후 1% 항-HIF-1α 항체[EPR16897 ](ab179483, Abcam, UK) 4℃에서 하룻밤. 그 후, 세포를 암실에서 1% CoraLite594 - 접합된 염소 항토끼 IgG(H + L)(Proteintech, USA)와 함께 1시간 동안 배양했습니다. 그런 다음 핵을 DAPI(Beyotime, Shanghai, China)로 염색하고 세포에 첨가하고 30초 동안 반응시켰다. 레이저 공초점 현미경(Olympus, Japan)을 사용하여 샘플을 검사했습니다. ImageJ v.1.4 분석 소프트웨어(Bethesda, MD, USA)를 사용하여 형광 강도를 정량화했습니다.
hWJ-MSC를 24시간 동안 CM(세포외 신호 조절 키나제(ERK)1/2, p-ERK1/2) 동안 배양하거나 골형성 배지에서 7일 동안 배양했습니다(runt-related transcription factor(RUNX)-2, type 1). 콜라겐/콜라겐 1(COL I)). 그런 다음, 세포를 RIPA 용해 완충액에서 30분 동안 용해했습니다. 세포 용해물을 원심분리하고 상층액을 분석을 위해 -20°C에서 보관했습니다. 12% SDS-PAGE 후, 단백질을 폴리비닐리덴 디플루오라이드(PVDF) 막으로 옮겼다. 사용된 1차 항체는 항-p-ERK1/2, 항-ERK1/2, 항-RUNX-2, 항-COL I 및 GAPDH(1:1,000, 토끼 다클론 항체; Cell Signaling Technology, Boston, MA, USA ). 결합되지 않은 항체를 제거한 후 막을 이차 항체와 함께 1시간 동안 배양했습니다. 막의 신호는 화학발광 젤 이미징 시스템(LAS4000M; GE Healthcare Biosciences AB, Uppsala, Sweden)을 사용하여 감지되었습니다. ERK에 대한 p-ERK 및 GAPDH에 대한 RUNX-2/COL I의 비율은 ImageJ v.1.4 분석 소프트웨어(Bethesda)를 사용하여 정량화되었습니다.
hWJ-MSC는 10 5 의 밀도로 시딩되었습니다. 6웰 플레이트의 웰당 세포. 부착 세포는 표시된 시간 동안 표시된 농도의 HIF-1α로 처리되었습니다. 그런 다음 세포를 수집하고 어둠 속에서 15분 동안 FITC-Annexin V 및 PI(Fcmacs, Nanjing, China)로 표지했습니다. 모든 샘플은 FACScan 유세포 분석기(BD Bioscience, San Jose, CA, USA)를 사용하여 테스트되었습니다. 데이터는 FlowJo v10(BD Biosciences)을 사용하여 분석되었습니다.
HAP의 엄청난 잠재력과 공동 배양 시스템 분석의 어려움을 고려하여 hWJ-의 골 형성 분화에서 HIF-1α의 역할을 이해하기 위해서는 정량적 분석과 신뢰할 수 있는 예측을 제공할 수 있는 수학적 모델이 필요했습니다. MSC.
hWJ-MSC는 0, 300, 500, 1000, 1500, 2000, 3000 및 4000pg/mL HIF-1α 및 골형성 유도액과 함께 배양되었습니다. 이 농도에서 데이터를 피팅한 후 대조군, m-HAP, np80 및 np20 그룹(각각 240, 300, 325 및 375pg/mL)에서 HIF-1α(HUVEC에서 생산)의 농도를 사용하여 MATLAB(MathWorks, Natick, MA, USA)을 사용하여 피팅 방정식을 테스트합니다. 모델을 단순화하기 위해 HIF-1α의 다른 농도에서 유사한 성장 패턴이 동일한 것으로 간주했습니다. 미분도-시간 방정식을 맞추기 위해 평균 미분도를 사용하였다. 다른 그룹에서 hWJ-MSC의 증식 속도, 세포 사멸 속도 및 골 형성 분화 정도가 정의 된 시간에 감지되었습니다.
실험 데이터에 따라 다단계 세포 계통 모델[18, 19]과 유사한 단순화된 2단계 세포 계통 모델이 수립되었습니다. C 0 및 C 1 hWJ-MSC 및 터미널 셀의 셀 번호를 각각 나타냅니다. C 0 및 C 1 다음에 의해 관리:
$$\left\{ \begin{gathered} \frac{{{\text{d}}C_{0} }}{{{\text{d}}t}} =\left[ {\frac{{K - C_{0} - C_{1} }}{K}p - (p - 1)} \right]\upsilon_{0} C_{0} \hfill \\ \frac{{{\text{d}} C_{1} }}{{{\text{d}}t}} =\left( {2 - \frac{{K - C_{0} - C_{1} }}{K}p - p} \ right)\upsilon_{0} C_{0} - AC_{1} \hfill \\ \end{gathered} \right.$$여기, p HIF-1α 및 시간의 영향을 받는 는 hWJ-MSC의 복제 확률을 나타냅니다. 이에 따라 d =1 − p 는 세포수의 추정 및 실험 데이터를 피팅하여 얻을 수 있는 분화율입니다. 매개변수 v0 세포가 각 계통 단계(특히 v =ln2/c , 여기서 c는 세포 주기의 지속 시간입니다. 말단 세포의 세포 사멸 속도는 A로 표시됩니다. . 간단하게 하기 위해 우리는 세포자멸사 비율이 시간이 지남에 따라 약간 변할 것이라는 점을 무시했으며 따라서 A =4.5%는 상수입니다. 케이 세포가 무제한 증식을 겪을 수 없기 때문에 환경 용량을 나타냅니다[20]. HIF-1α는 hWJ-MSC의 분화율을 증가시켜 다음과 같은 분화율을 나타냅니다.
$$\begin{정렬} d &=\frac{{d_{0} }}{{1 + (r*H)^{m} }} \\ p &=1 - d \\ \end{정렬} $$여기서 H HIF-1α의 농도를 나타내고; d 0 0pg/mL HIF-1α에서의 분화율을 나타냅니다. r은 조절의 강도를 나타내고(여기서, MSC에 대한 HIF-1α의 조절의 강도를 나타냄); m은 힐 계수[21]에 해당하며 MSC 분화율과 HIF-1α 농도 사이의 관계를 설명합니다.
정규성 및 등분산성 요구 사항을 충족하는 모든 데이터는 3개 이상의 독립적인 실험에서 평균 ± 표준 편차(SD)로 표시됩니다. SPSS 24.0 소프트웨어(SPSS Inc., Chicago, USA)를 사용하여 일원 ANOVA 또는 양방향 ANONA를 통한 통계 분석을 수행했습니다. A P 값 <0.05는 통계적으로 유의한 것으로 간주되었습니다. 통계 분석은 GraphPad Prism 5(GraphPad Software, La Jolla, CA)를 사용하여 제공됩니다.
<섹션 데이터-제목="결과">그림 2와 같이 특정 크기와 모양으로 HAP를 준비했습니다. 구형에 가까운 np20의 직경은 평균 20nm였고, np80은 평균 길이 80nm, 너비 20nm로 막대 모양이었습니다. m-HAP는 또한 모양이 거의 구형이고 직경이 약 12μm였습니다. 모든 입자는 L-DMEM에서 음의 표면 전하를 가졌습니다. 제타 전위의 음의 값은 뼈 세포의 부착 및 증식뿐만 아니라 직접적인 뼈 결합 및 새로운 뼈 형성에 상당한 유리한 영향을 미친다고 제안되었습니다[22, 23]. L-DMEM에서 관찰된 입자는 수성 시스템에서 응집하는 경향이 있습니다. 그들의 유체역학적 크기도 테스트되었는데, 이는 생물학적 행동에 영향을 미치는 중요한 요인일 수도 있습니다.
<그림>
HAP의 특성화. a의 TEM 현미경 사진 np20 및 b np80 및 c의 SEM 현미경 사진 m-HAP. d HAP의 특성화(n =6). 약어:TEM 투과 전자 현미경, SEM 주사 전자 현미경, HA 수산화인회석, m-HAP 마이크로 크기의 HAP 입자
hWJ-MSC에 대한 HAP의 간접 독성을 평가하기 위해 MTS 분석을 통해 세포 생존력을 측정했습니다. 50µg/mL HNP가 포함된 CM은 24시간 및 72시간 후, 특히 24시간 후에 hWJ-MSC 생존력을 크게 자극할 수 있습니다. 그러나 100µg/mL HANP가 포함된 CM은 72시간 후 대조군에 비해 세포 생존율을 15~20% 감소시켰습니다. 또한 np80이 아닌 25µg/mL np20을 포함하는 CM은 24시간 후 세포 생존을 자극했습니다. 이러한 현상은 50 µg/mL HAP가 모든 후속 실험에서 사용된 세포독성 농도 이하임을 확인시켜줍니다(그림 3).
<그림>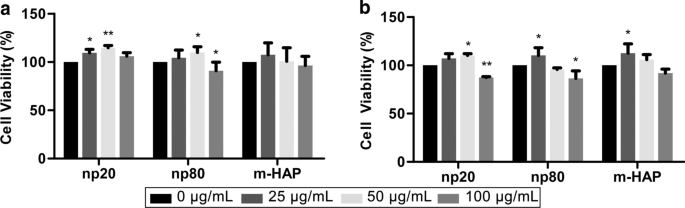
hWJ-MSC에 대한 HAP의 간접 독성. HAP와 간접적으로 배양된 hWJ-MSC의 생존력은 a에 대해 측정되었습니다. 24 및 b 72시간 *피 <0.05; **피 <0.01 대 대조군. 대조군은 HAP 처리 없이 CM에서 배양된 세포로 구성되었으며 세포 생존율은 대조군의 백분율로 정규화되었습니다. 약어:HAP 수산화인회석 입자, m-HAP 마이크로 크기의 HAP 입자, hWJ-MSC 인간 탯줄 와튼 젤리 유래 중간엽 줄기세포
hWJ-MSC에 대한 HANP의 간접적인 골유도 효과를 확인하기 위해, 골형성 유전자의 발현을 정량적 RT-PCR 분석에 의해 평가하였다. HANP 그룹, 특히 np20에서 런트 관련 전사 인자 2(RUNX-2)의 전사 수준은 7일에서 21일까지 눈에 띄게 증가했습니다(그림 4a). np20 그룹에서 I형 콜라겐(Col I)의 유전자 발현은 7일째부터 21일째까지 향상을 입증한 반면, np80 그룹은 7일째부터 14일째까지 지속적인 증가를 보여주었습니다(그림 4b). Osteocalcin(OCN) mRNA는 14일째에 HANP 그룹에서 분명히 상향 조절되어 골형성의 가속화된 속도를 나타냅니다(그림 4c). ALP(alkaline phosphatase)의 mRNA 수준은 HANP 그룹에서 분명히 증가했으며(그림 4d), 이는 hWJ-MSC의 골 형성 분화를 나타냅니다. 그러나 m-HAP군은 대조군에 비해 이들 3가지 골형성 유전자의 수준에 있어 제한적인 변화를 보였다(Fig. 4). 또한, 만능 마커인 NANOG, OCT3/4 및 SOX2의 발현은 대조군과 비교하여 HANP 그룹에서 감소했으며(그림 4e-g), 이는 HANP 그룹의 hWJ-MSC가 특히 다음에서 분화되었음을 의미합니다. np20 그룹. Western blot analysis(그림 5c, d)에서도 유사한 결과를 얻었으며, 이는 np 20 그룹이 hWJ-MSC에서 RUNX-2 및 COL I의 발현을 간접적으로 향상시킬 수 있음을 나타냅니다.
<그림>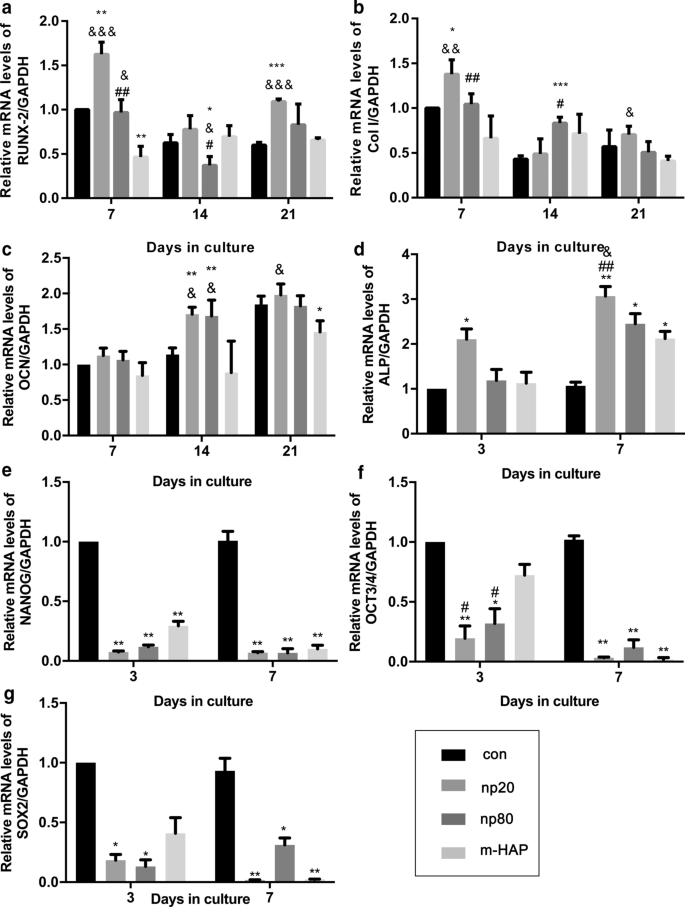
골 형성 분화 관련 유전자의 발현에 대한 HAP의 간접 효과. 아 RUNX-2, b Col I, c OCN, d ALP, e 나노그, f 10월 3/4, g 표시된 시간 동안 CM과 함께 배양된 hWJ-MSC의 SOX2 유전자 수준. *피 <0.05; **피 <0.01; ***피 <0.001 대 대조군; & 피 <0.05; && 피 <0.01; &&& 피 <0.001 대 m-HAP 그룹; # 피 <0.05; ## 피 <0.01 대 np20 그룹. HAP를 처리하지 않은 골형성 배지에서 배양한 세포를 대조군으로 사용하였다. 약어:HAP 수산화인회석 입자, m-HAP 마이크로 크기의 HAP 입자, hWJ-MSC 인간 탯줄 와튼 젤리 유래 중간엽 줄기세포 RUNX-2 런트 관련 전사 인자 2, Col I I형 콜라겐, OCN 오스테오칼신, ALP 알칼리성 인산분해효소, SOX 2 SRY 관련 HMG-box 2
<그림>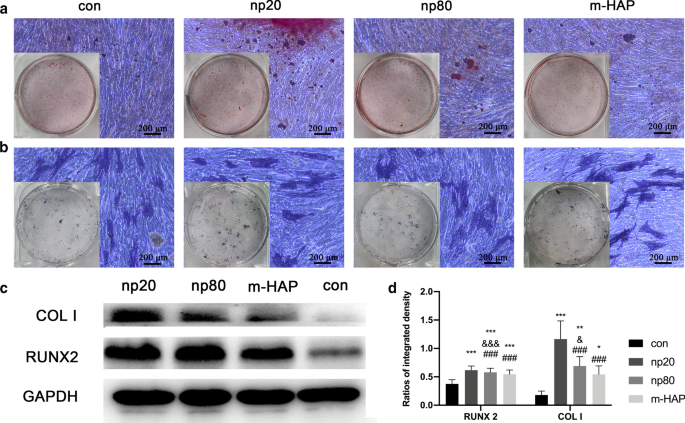
세포 외 칼슘 침착 및 ALP 활성에 대한 HAP의 간접 효과. hWJ-MSC를 14일 동안 골형성 배지에서 배양하였다. 아 세포 외 칼슘 침착은 ARS 염색을 통해 시각화되었습니다. ㄴ hWJ-MSC의 ALP 활성은 ALP 염색, 눈금 막대:200μm를 통해 평가되었습니다. ㄷ 웨스턴 블롯 분석은 7일째에 골형성 배지에서 hWJ-MSC의 RUNX-2 및 COL I의 발현을 나타내었습니다. d 부품(c)에서 RUNX-2 및 COL I의 농도 측정 ). HAP를 처리하지 않은 골형성 배지에서 배양한 세포를 대조군으로 사용하였다. *피 <0.05; **피 <0.01; ***피 <0.001 대 대조군; & 피 <0.05; &&& 피 <0.001 대 m-HAP 그룹; ### 피 <0.001 대 np20 그룹. 약어:HAP 수산화인회석 입자, m-HAP 마이크로 크기의 HAP 입자, hWJ-MSC 인간 탯줄 와튼 젤리 유래 중간엽 줄기세포, ALP 알칼리성 인산분해효소
hWJ-MSC에 대한 HANP의 간접적인 골유도 효과를 시각적으로 관찰하기 위해 세포를 표시된 골형성 배지와 함께 14일 동안 배양한 후 ARS 및 ALP 염색을 하였다. 도 5a, b에 나타난 바와 같이 m-HAP 및 대조군에 비해 HANP군에서 광물화된 결절의 수가 증가하고 hWJ-MSC의 높은 ALP 활성이 관찰되었다. 또한, 대조군과 유사한 m-HAP는 hWJ-MSC의 골 형성 분화에 제한된 효과를 보였습니다.
HAP에 의한 HUVEC의 측분비 기능에 대한 영향을 조사하기 위해 면역형광 및 ELISA 분석을 사용하여 hWJ-MSC의 골형성 분화를 촉진하는 가능한 단백질을 확인했습니다. 그림 6a-c에서 볼 수 있듯이 HIF-1α의 세포 내 및 세포 외 생산은 HAP(특히 np20)에 의해 크게 촉진되었지만 m-HAP의 생산에 대한 영향은 제한적이었습니다.
<그림>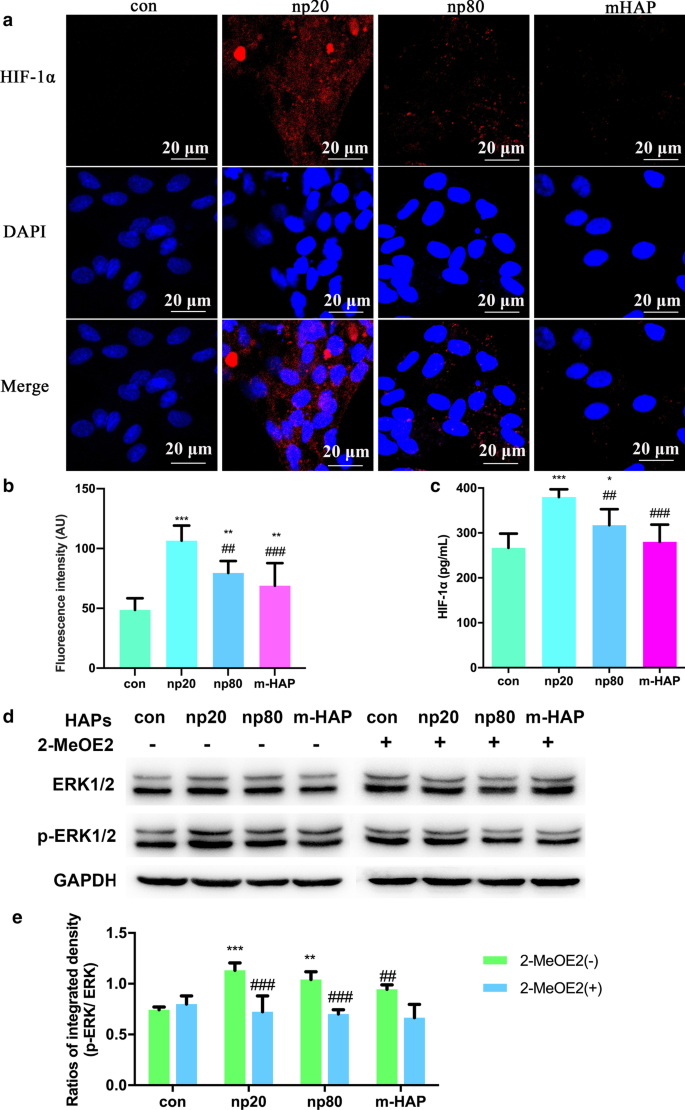
HAP는 HUVEC와 간접적으로 공동 배양된 hWJ-MSC에서 ERK1/2 신호 전달을 활성화했습니다. 아 18시간 동안 HAP가 있거나 없는 HUVEC에서 수행된 HIF-1α의 면역형광. ㄴ 부분(a)에서 HIF-1α의 형광 강도 ). ㄷ 18시간 동안 HAP를 처리하거나 처리하지 않은 HUVEC의 배양 배지에서 HIF-1α의 세포외 농도를 ELISA를 통해 측정했습니다. 스케일 바:20μm. *피 <0.05; **피 <0.01; ***피 <0.001 대 대조군; ## 피 <0.01; ### 피 <0.001 대 np20 그룹. HAP를 처리하지 않은 세포를 대조군으로 사용하였다. hWJ-MSC는 24시간 동안 CM으로 처리되었습니다. d ERK1/2 경로에서 주요 키나제의 활성화를 나타내는 웨스턴 블롯 분석. 이 부품(b)에서 p-ERK1/2의 밀도 측정 ). **피 <0.01; ***피 <0.001 대 대조군; ## 피 <0.01, ### 피 <0.001 대 2-MeOE2( −) 그룹. HAP 처리 없이 CM에서 배양된 세포를 대조군으로 사용하였다. 약어:HAP 수산화인회석 입자, m-HAP 마이크로 크기의 HAP 입자, hWJ-MSC 인간 탯줄 Wharton의 젤리 유래 중간엽 줄기 세포, HUVEC 인간 제대 정맥 내피 세포, ERK 세포외 신호 조절 키나제, HIF-1α 저산소증 유발 인자 1α
HIF-1α에 의해 활성화된 hWJ-MSC의 분화 신호 전달 경로를 보다 정확하게 이해하기 위해 우리는 웨스턴 블롯 분석을 통해 ERK1/2 경로의 주요 조절자를 조사했습니다. 그림 6d, e에서 볼 수 있듯이 총 ERK1/2의 단백질 수준은 변경되지 않은 상태로 유지되었지만 p-ERK1/2 수준은 HANP와 함께 배양된 hWJ-MSC에서 뚜렷하게 증가했으며 이는 np20 그룹에서 특히 그렇습니다. 그러나 m-HAP는 HUVEC에서 HIF-1α 생산에 미치는 영향과 유사하게 hWJ-MSC에서 p-ERK1/2 수준에 거의 영향을 미치지 않았습니다. 중요하게도, HIF-1α에 의해 활성화된 hWJ-MSC에서 증가된 수준의 p-ERK1/2는 특정 HIF-1α 억제제인 2-MeOE2에 의해 차단될 수 있으며, 이는 HIF-1α가 ERK1/2 신호 전달 경로의 상류에서 기능했음을 나타냅니다. hWJ-MSC에서.
관찰된 hWJ-MSC 골형성 분화 자극에 HIF-1α가 필요한지 여부를 결정하기 위해 특정 HIF-1α 억제제(2-MeOE2)를 이러한 세포 배양물에 적용했습니다. 도 7에 나타난 바와 같이, 골형성 배지에서 배양된 2-MeOE2(+) 처리군의 hWJ-MSCs의 광물화된 기질 침착 및 ALP 활성이 약화되었고, 이는 HIF-1α가 hWJ-의 골형성 분화에 필수적임을 시사한다. MSC. 이러한 결과를 바탕으로 우리는 HIF-1α에 의해 활성화된 hWJ-MSC의 골 형성 분화에서 ERK1/2 경로의 역할을 추가로 조사했습니다. 특정 MEK 억제제인 PD98059 투여 후 골형성 배지로 배양된 hWJ-MSC에서 광물화된 기질 침착 및 ALP 활성이 억제되었습니다.
<그림>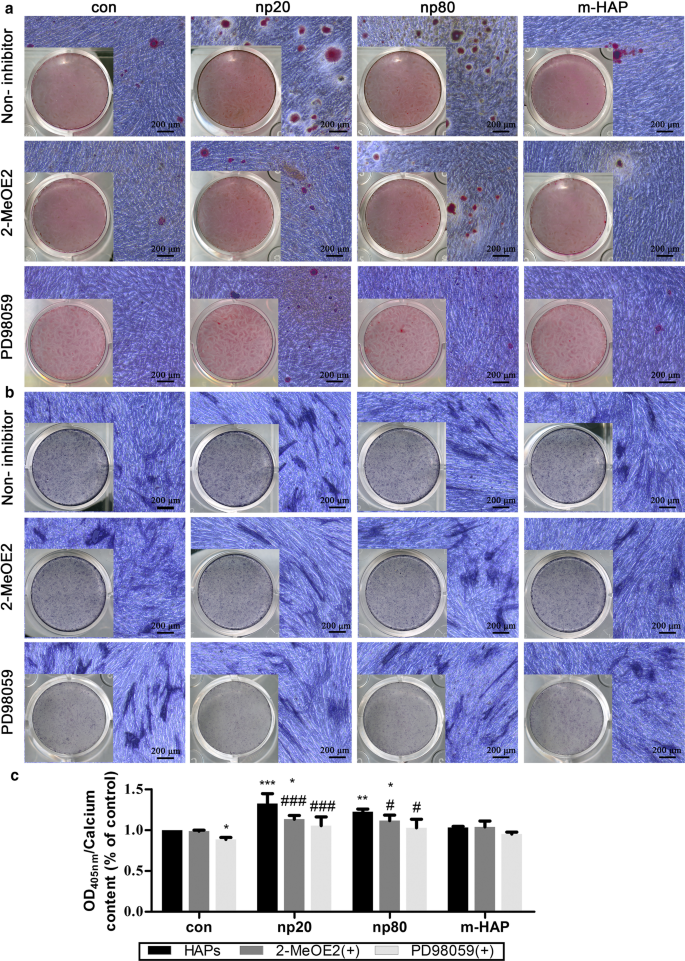
HIF-1α는 ERK1/2 경로를 통해 hWJ-MSC의 골형성 분화를 촉진했습니다. hWJ-MSC를 14일 동안 PD98059가 있거나 없는 골형성 배지에서 배양했습니다. 아 세포외 칼슘 침착은 ARS 염색을 통해 시각화되었습니다. ㄴ hWJ-MSC의 ALP 활성은 ALP 염색을 통해 평가되었습니다. 스케일 바:200μm ㄷ 세포외 칼슘 기질의 정량 분석. *피 <0.05; **피 <0.01; ***피 <0.001 대 대조군; # 피 <0.05; ### 피 <0.001 대 np20 그룹. Cells incubated in osteogenic medium without HAPs and PD98059 treatment were used as the control group. Abbreviations:HAPs hydroxyapatite particles, m-HAP micro-sized HAP particles, hWJ-MSCs human umbilical cord Wharton’s jelly-derived mesenchymal stem cells, HUVECs human umbilical vein endothelial cells, ERK extracellular signal-regulated kinase, HIF-1α hypoxia-inducible factor 1α, ALP alkaline phosphatase
To quantitatively reveal the intrinsic connection between the concentration of HIF and osteogenic differentiation of hWJ-MSCs, 0–4000 pg/mL of HIF-1α was used to treat eight groups of hWJ-MSCs. A two-stage cell-lineage mathematical model was used to analyze the proliferation, apoptosis, and osteogenic differentiation rates of these hWJ-MSCs treated with different concentrations of HIF-1α. As shown in Fig. 8a, fitting data were employed to obtain the simulated formula (\(\frac{d}{{d_{0} }} =\frac{1}{{0.14H^{2} - 0.43H + 1}}\)) and curve (blue curve), showing that the differentiation rate first increased and then decreased with the increase in HIF concentration. More specifically, the differentiation rate reached a peak at 1500 pg/mL HIF-1α.

Two-stage cell-lineage models. hWJ-MSCs were incubated in a defined concentration of HIF-1α for the indicated times. 아 Relative differentiation rates of hWJ-MSCs at different concentrations of HIF-1α, b relative ALP activity (differentiation degrees of hWJ-MSCs) and relative osteoblast cells on different culture days. ㄷ Three-dimensional surface of differentiation degree evolving with time and HIF-1α. Cell number with an initial seeding density of 1500 cells per well (in a 96-well plate), as well as d 0, e 375, and f 1500 pg/mL HIF-1α. Abbreviations:hWJ-MSCs human umbilical cord Wharton’s jelly-derived mesenchymal stem cells, HIF-1α hypoxia-inducible factor 1α, ALP alkaline phosphatase
According to our study, the concentrations of HIF-1α produced by HUVECs in the control, m-HAP, np80, and np20 groups were 240, 300, 325, and 375 pg/mL, respectively. The black square represents the differentiation rate promoted by 240, 300, 325, and 375 pg/mL HIF-1α, and this matches well with the simulated curve. In addition, the differentiation degrees of the hWJ-MSCs (relative ALP activity) treated with different concentrations of HIF-1α increased similarly increased with time. Therefore, in order to simplify the model, we consider their growth patterns to be similar or even identical. We found that the increase in ALP activity from Day 0 to Day 7 was proportional to the number of osteoblasts, and osteoblast cells reached their peak at the platform period. Therefore, after fitting the ALP activity with the relative osteoblasts cell number curve (osteoblasts cell number / maximum osteoblasts cell number), the maximum of ALP activity was predicted and relative ALP activity (ALP activity/ maximum of ALP activity), representing the differentiation degree, was acquired (Fig. 8b). Combining these two studies, the three-dimensional surface of the differentiation degree evolving with time and HIF-1α was obtained (Fig. 8c).
In order to estimate the optimal culture time, it was necessary to simulate cell numbers. After elucidating the differentiation rate under different concentrations of HIF-1α, we simulated the size of each population. The simulation utilized an initial seeding density of 1500 cells per well (in a 96-well plate) using different concentrations of HIF-1α. As shown in Fig. 8d–f, the experimental data (black square) match well with the simulated total cell numbers (blue curve), which is a sum of the number of hWJ-MSCs and osteoblasts, and this supports the ability of this model to predict the number of hWJ-MSCs and osteoblasts at any time point. Moreover, the osteoblast cell number approached the platform period at 21, 18, and 15 days with 0, 375, and 1500 pg/mL HIF-1α, respectively. This model provides the optimum culture time for guiding tissue engineering, and it also provides direct evidence that HIF-1α accelerates the osteogenic differentiation of hWJ-MSCs.
With recent advances in nanobiomaterials, nano-based artificial bone substitutes have been an area of intense investigation. The accumulating evidence suggests that there are complex interactions between cells and nanobiomaterials due to their capacity to penetrate cell membranes and increase internal retention times [24, 25]. A previous study revealed that collagen/alginate nanofilms can adsorb onto the MSC membrane to activate intracellular signaling cascades and promote osteogenic differentiation [26]. Elegant experiments by Wu and his colleagues clearly demonstrated that TiO2 nanotubes can improve vascularization and osteogenic differentiation by facilitating paracrine effects and cell junctions via EC-MSC interactions [27]. For the purpose of developing excellent candidates for bone tissue engineering, it is necessary to clarify the direct crosstalk between nano-based bone substitutes and cells implicated in bone repair as well as their indirect interactions. However, our current understanding of this is still limited. In the present study, we utilized an indirect co-culture model to further elucidate the biological effects of HANPs on MSCs in regard to the indirect interactions mediated by ECs.
Cytotoxicity is a primary issue for assessing the biocompatibility of any nanobiomaterial. Although our previous study found that HANPs did not directly influence the viability or apoptosis of hWJ-MSCs, they may still exert different impacts via the mediation of other cells [28]. Thus, it was necessary to evaluate the cytotoxic effects of HANPs on hWJ-MSCs mediated by HUVECs. Interestingly, after incubation in CM for 24 h and 72 h, hWJ-MSC viability was maintained and even elevated in the 0–50 µg/mL HANP groups, especially in the np20 group, indicating the existence of effector molecules in the CM. When the concentration of HANPs reached 100 µg/mL, they became cytotoxic to the hWJ-MSCs. However, 0–100 µg/mL m-HAP had no influence on hWJ-MSC viability (Fig. 3). Jiang et al. have shown that engineered nanoparticles of a particular size can have distinct endocytic routes and kinetics associated with altered downstream signaling involved in regulating target cell functions [29]. In our previous study, we showed that np20 and np80 were endocytosed by HUVECs, and this was followed by morphologic changes and the appearance of large vacuoles, indicating the activated state of the HUVECs. Additionally, np20, with their faster uptake speed and increased accumulation, might result in a stronger activation of HUVECs, possibly resulting in increased hWJ-MSC viability via paracrine signaling. Conversely, few m-HAPs can be endocytosed by HUVECs, and this might account for their limited influence on the metabolism of hWJ-MSCs [9].
To further explore the potential osteoinductive effect of activated HUVECs, a subcytotoxic dose of 50 µg/mL HAPs was used in subsequent studies. The CM collected from the activated HUVECs promoted extracellular calcium deposition, ALP activity, and osteogenic proteins expression in hWJ-MSCs, as well as the mRNA expression of osteogenic genes (Figs. 4, 5). Runx2, an essential transcription factor involved in specifying the osteoblast lineage [30], showed a substantial enhancement in the np20 group, indicating a strong osteoinductive effect on hWJ-MSCs (Fig. 4a and Fig. 5c, d). The np20 group demonstrated a 1.5-fold improvement in COLI expression at Day 7 (Figs. 4b, 5c, d) and a double increase at Day 14, which implied the presence of additional differentiated osteoblasts in the HANP-treated groups (Fig. 4b) [30]. OCN is a mature stage bone marker [31], and this gene showed a significant increase in the HANP groups at Day 14 (Fig. 4c), indicating that np20 and np80 can accelerate bone maturation compared to m-HAP. ALP is an early marker of osteoblast differentiation, and it obviously increased with culture time in each group, especially the np20 group, revealing that additional transformation occurred from MSCs to osteoblasts (Fig. 4d). Pluripotency markers, NANOG, OCT3/4, and SOX2 imply the capacity for differentiation [32]. As shown in Fig. 4e–g, the decreased expression in the genes of the HANP groups implied that most of the hWJ-MSCs in HAP groups had transformed into osteoblasts.
Our data demonstrated that the endocytosis of HANPs by HUVECs was associated with an improved osteogenic differentiation of hWJ-MSCs. However, the cause of this outcome is currently unclear. In terms of the paracrine function of HUVECs, we focused on soluble differentiation-inducing proteins in the supernatant of activated HUVECs. HIF-1α signaling is essential in coupling ossification and angiogenesis during bone regeneration [33, 34]. Heikal et al. reported that injured ECs secrete more HIF-1α even under normoxia conditions [35]. It has also been shown that exposure to HANPs inhibits the angiogenic ability of HUVECs [9]. Thus, we measured the concentrations of HIF-1α in the CM, and the results showed that the HIF-1α content increased in the HANP treatment groups compared to the m-HAP and control groups (Fig. 6a). To identify the role of HIF-1α in the osteogenic differentiation of hWJ-MSCs, we used 2-MeOE2, which is a specific HIF-1α inhibitor, was used. The decreased concentration of HIF-1α paralleled the impaired mineralized matrix deposition and ALP activity in these hWJ-MSCs, indicating that HANPs can promote the HIF-1α production of HUVECs to facilitate the osteogenesis of hWJ-MSCs (Fig. 7).
To properly apply HANPs for use in bone tissue engineering, it is necessary to gain further insights into the mechanisms by which HANPs promote the osteogenic differentiation of hWJ-MSCs mediated by HUVECs. The ERK1/2 pathway is downstream of HIF-1α [36] and is fundamental to the differentiation of MSCs [37]. In this work, the concentrations of HIF-1α in the CM coincide well with the p-ERK1/2 levels in the hWJ-MSCs (Fig. 6b, c). When 2-MeOE2 was applied, the p-ERK1/2 expression in the hWJ-MSCs failed to be activated, indicating that HIF-1α functioned upstream of ERK1/2 signaling. To directly address the role of ERK1/2 signaling in the osteogenic differentiation of hWJ-MSCs, PD98059, a specific MEK inhibitor, was used. The suppression of ERK1/2 signaling resulted in the lowest osteogenic differentiation of hWJ-MSCs. One possible reason for this occurrence is that the ERK1/2 pathway plays a key role in both HIF-1α signaling and in the apoptosis and proliferation signaling pathways, which could be responsible for the observed changes in osteogenic differentiation in these cells [38, 39]. Additionally, this could also be related to the presence of vascular endothelial growth factor (VEGF). VEGF is one of the downstream effectors of HIF-1α signaling [33], and it can also promote the osteogenic differentiation of MSCs via activation of the ERK1/2 pathway [37]. Our previous study found that np20 induced the production of VEGF in HUVECs [9]; therefore, it is possible that the suppression of the ERK1/2 pathway may result in inhibition of VEGF, which would lead to the decreased osteogenic differentiation of hWJ-MSCs. According to the available experimental results, we can summarize as follows. HANPs are able to more optimally process better direct [5] and indirect osteoinductive effects than m-HAPs. Compared to autogenous bone grafts and bone allografts, there is an extensive source of HANP and without secondary damage and potential immunogenicity. However, compared to m-HAPs, HANPs can suppress the angiogenic ability of HUVECs [9] and exhibit slight cytotoxicity in both a time- and dose-dependent fashion.
Recently, growing evidence has demonstrated the importance of HIF-1α in the bone regeneration. However, few studies have been able to quantitatively predict the MSC differentiation rate under specific initial conditions, such as the HIF-1α concentration. Taking cell proliferation, apoptosis, and osteogenic differentiation into account, we present a mathematical model that combines a two-stage cell lineage with HIF-1α that is highly correlated with our experimental data. By fitting the differentiation rate of hWJ-MSCs in 0–4000 pg/mL HIF-1α, we acquired the equations for describing the differentiation rate, HIF-1α concentration, and time. As shown in Fig. 8d, this model can depict the cell number map under different HIF-1α concentrations, so that it is possible to explore the intrinsic dynamics of the two-stage system [40]. Additionally, this model mathematically validates the effect of HIF-1α on the osteogenic differentiation of hWJ-MSCs. Moreover, based on a multi-stage cell-lineage model and logistic model, our model is sufficiently stable to enable long-term predictions without falling into the trap of population unlimited explosion [41].
By using the existing experimental data, both the cell number and differentiation rate can be predicted with a defined initial cell seeding density and HIF-1α concentration. As such, the optimum incubation time is also obtained. Consequently, we can predict the optimum concentration of HIF-1α and determine the most optimal time for osteogenesis, which is important for efficient tissue engineering. A two-stage cell-lineage model is applicable for predicting the proliferation and differentiation of stem cells, which have two cell lineages. On this basis, the model founded on the initial conditions and existing experimental data can be established to identify the optimum culture conditions in vitro, which will assist in optimizing bone repair in vivo.
In this study, we explored the specific biological effects of HANPs on hWJ-MSCs mediated by HUVECs. Compared to m-HAPs, both np20 and np80 showed slight cytotoxicity in both a time- and dose-dependent fashion. Importantly, the size of the HANPs appeared to have no significant impact on this cytotoxicity. Our data also showed that HANPs, especially np20, were capable of facilitating HUVECs to secrete increased levels of HIF-1α, which directly correlated with the enhanced osteogenic differentiation of hWJ-MSCs via the activation of the ERK1/2 pathway (Fig. 9). More remarkably, the results from the two-stage cell-lineage model suggested that HIF-1α exerted a dose-dependent stimulatory effect on the osteogenic differentiation rate of hWJ-MSCs. Additionally, the optimum concentration of HIF-1α and incubation time were estimated based on the initial conditions using an in vitro model, which could be invaluable in the future for tissue engineering applications. Collectively, these observations provide evidence that HANPs may improve bone regeneration by modulating cell–cell interactions.

A schematic illustration of the possible mechanisms
현재 연구 중에 사용 및 분석된 데이터 세트는 합당한 요청이 있는 경우 교신저자에게 제공됩니다.
Hydroxyapatite
HA nanoparticles
Human umbilical cord Wharton’s jelly-derived mesenchymal stem cells
인간 제대 정맥 내피 세포
Micro-sized HAP particles
인산염 완충 식염수
EC medium
2-Methoxyestradiol
효소 결합 면역흡착 분석
Runt-related transcription factor 2
Type I collagen
Osteocalcin
알칼리성 인산분해효소
SRY-related HMG-box 2
Extracellular signal-related kinases
Vascular endothelial growth factor
나노물질
초록 심근염은 효과적인 치료 없이 심근의 국소화 또는 확산 염증을 특징으로 하는 질병이다. 이 연구는 조절을 통해 바이러스성 심근염(VMC) 쥐의 심근 섬유증 및 상피-중간엽 전이(EMT)에 대한 골수 중간엽 줄기세포 유래 엑소좀(BMSC-Exo)에서 분비되는 microRNA-133(miR-133)의 조절 메커니즘을 탐구했습니다. 마스터마인드 1(MAML1). 쥐의 BMSC를 분리 배양하여 면역 표현형과 골 형성 및 지방 형성 능력을 확인하고 BMSC-Exo를 추출하여 확인했습니다. 초원심분리를 통해 엑소좀을 얻었고, 투과전자현미
초록 파킨슨병(PD)은 중뇌에서 도파민성 뉴런의 점진적인 소실을 특징으로 하며, 줄기 세포 이식 방법은 치료를 위한 유망한 전략을 제공합니다. 이러한 연구에서 생체 내 이식된 세포의 생물학적 행동을 추적하는 것은 줄기 세포 기능에 대한 기본 이해와 임상 효과 평가에 필수적입니다. 본 연구에서 우리는 열분해 방법과 2단계 개질을 통해 폴리아크릴산(PAA)과 메톡시폴리에틸렌글리콜아민(PEG)으로 코팅된 새로운 초소형 초상자성 산화철 나노입자를 개발했습니다. USPIO-PAA/PEG NP는 TEM 및 FTIR에서 볼 수 있듯이 10.0