

나노물질

산업 제조
microRNA를 발현하는 인간 탯줄 중간엽 줄기 세포(hucMSC)에서 유래한 엑소좀은 인간 질병에서 강조되었습니다. 그러나 자간전증(PE)에 대한 hucMSCs 유래 엑소좀 miR-18b-3p의 자세한 분자 메커니즘은 추가 조사로 남아 있습니다. 우리는 엑소좀과 miR-18b-3p/렙틴(LEP)이 PE의 발생에 미치는 영향을 조사하는 것을 목표로 했습니다. hucMSC 및 hucMSC-엑소좀(Exos)의 형태를 확인하였다. 엑소좀은 PE에서 miR-18b-3p의 역할을 탐구하기 위해 miR-18b-3p를 발현하는 다른 렌티바이러스에 감염되었습니다. PE 래트 모델은 N의 복강내 주사에 의해 확립되었습니다. -니트로-1-아르기닌 메틸 에스테르. LEP 및 miR-18b-3p의 발현은 PE 래트 태반 조직에서 테스트되었습니다. 또한, LEP 및 miR-18b-3p 발현에 대한 엑소좀의 효과가 검출되었다. 수축기 혈압(SBP), 단백뇨, 염증 인자, 태아 쥐와 태반의 무게, PE 쥐의 세포 사멸이 감지되었습니다. 마지막으로 이중 루시퍼라제 리포터 유전자 분석과 RNA 풀다운 분석을 사용하여 miR-18b-3p와 LEP 사이의 관계를 확인했습니다. miR-18b-3p를 복원하거나 LEP를 억제하는 엑소좀은 PE 쥐의 SBP와 단백뇨를 감소시켰을 뿐만 아니라 태아 쥐와 태반의 무게를 증가시켰고, 염증 인자의 혈청 수준을 감소시켰을 뿐만 아니라 PE 쥐의 세포 사멸 세포를 억제하여 억제 효과를 발휘했습니다. PE 진행에 대해. PE 쥐의 태반 조직에서 miR-18b-3p가 감소하고 LEP가 증가했습니다. LEP는 miR-18b-3p의 직접적인 표적 유전자였다. miR-18b-3p의 상향 조절 또는 엑소좀 처리는 LEP 발현을 억제하고 PE 발생을 감소시킨 반면 miR-18b-3p의 하향 조절은 반대 효과를 나타냈다. 하향 조절된 LEP는 PE 쥐에 대한 miR-18b-3p 감소 효과를 역전시켰습니다. HucMSCs 유래 엑소좀 miR-18b-3p는 LEP가 PE의 발생 및 발달에 참여하도록 표적화합니다. 이 연구는 PE의 메커니즘과 조사에 대한 새로운 이론적 기초를 제공할 수 있습니다.
단백뇨와 고혈압을 특징으로 하는 자간전증(PE)은[1] 인간 임신에서 태아 및 산모 사망 및 이환율의 주요 원인입니다[2]. PE의 병인 및 병인은 명확하지 않으며[3], 비정상적인 영양막 침윤으로 인해 모체 내피 기능 장애, 만성 태반 관류 이상 및 고혈압을 유발하는 부작용이 있는 것으로 보고되었습니다[4]. 태아 및 태반 분만을 제외하고 PE에 대한 특정 치료법은 없습니다[5]. 따라서 이 질병의 예후를 개선하기 위한 치료 목표를 찾는 것이 시급합니다.
인간 탯줄(huc)은 다양한 영양 인자와 사이토카인을 분비할 뿐만 아니라 강력한 항염증 및 면역 조절 능력을 나타내는 중간엽 줄기 세포(MSC)의 적합한 공급원입니다[6]. 연구에 따르면 PE가 탯줄 Wharton의 젤리 유래 MSC에서 신경교 마커 발현을 가속화한다는 것이 확인되었습니다[7]. PE 쥐의 태반 형태와 혈관신생에 대한 hucMSC 엑소솜(Exos)의 보호 효과도 보고되었습니다[8]. 엑소좀은 작은(50-100nm) 분비 소포로, 순환에 의해 암 유발 인자를 캡슐화하고 먼 부위나 주변 세포로 전달함으로써 종양 미세 환경에서 세포 간의 의사 소통을 매개합니다[9]. 한 연구에 따르면 엑소좀에 의한 PE 환자에서 sFlt-1 및 sEng의 내피 세포로의 효과적인 전달에 의해 유발된 혈관 기능의 손상 및 합병증이 밝혀졌습니다[10]. MicroRNA(miRNA)는 길이가 18-25 뉴클레오티드인 내인성 비암호화 RNA이며 전사 후 수준에서 유전자 발현을 조절합니다[11]. 연구의 데이터는 miR-18b 발현이 PE에서 세포 침윤, 생존력 및 영양막 세포의 이동에 영향을 미친다고 보고했습니다[12]. 또한, Wu et al. miR-18b가 고포도당에 의해 유도된 인간 망막 내피 세포의 증식을 약화시키며, 이는 당뇨병성 망막증 발병 기전을 이해하는 데 새로운 통찰력을 제공할 수 있다고 제안했습니다[13]. 그러나 PE에서 hucMSC 유래 엑소좀 miR-18b-3p의 역할은 아직 알려지지 않았습니다. 렙틴(LEP)은 생리학적 상태의 세포 분화/증식 및 면역에 다면발현 효과를 가지며 주로 태반을 포함한 다른 조직 외에 지방세포에서 나온다[14]. 연구에 따르면 비정상적인 LEP 프로모터 메틸화가 PE 진행에 관여한다는 것이 확인되었습니다[15]. 또 다른 연구에서는 태반이 임신 중 LEP 발현의 주요 부위라고 제안했습니다[16]. 그럼에도 불구하고 miR-18b-3p와 LEP 사이의 결합 관계는 여전히 애매합니다. 따라서 우리는 LEP와 함께 PE에서 hucMSC 유래 엑소좀 miR-18b-3p의 역할을 탐구하는 것을 목표로 했으며, hucMSC 유래 엑소좀 miR-18b-3p가 LEP를 표적으로 하여 PE 진행을 억제할 수 있다고 추론했습니다.
이 연구는 우한대학교 인민병원 기관심사위원회의 승인을 받았습니다. 모든 참가자는 사전 동의 문서에 서명했습니다. 모든 동물 실험은 우한 대학 인민 병원 국제 위원회의 실험 동물 관리 및 사용 지침에 따라 집계되었습니다.
건강한 갱년기가 낳은 태아의 탯줄을 채취하여 잘게 썰고 체로 걸러낸 다음 PBS(phosphate-buffered saline) 용액과 섞습니다. 탯줄 조직을 10cm의 원심 반경으로 5분 동안 1500r/min의 속도로 원심분리했습니다. 조직을 10% 소태아혈청(FBS)을 함유하는 둘베코의 변형된 독수리 배지(DMEM)/F12로 현탁시키고 배양 플라스크로 옮겼다. 액체는 4일 후에 교체한 다음 3일마다 한 번씩 교체했습니다. 합류가 약 90%에 도달했을 때 세포를 계대배양하였다. 광학현미경으로 hucMSCs의 부착성 성장과 형태를 관찰하였다. 세포를 oil red O 염색 용액(Beyotime Institute of Biotechnology, Shanghai, China)으로 염색하여 hucMSC의 골형성 분화를 검출하고, 알칼라인 포스파타제(ALP) 염색 용액(Beyotime)으로 염색하여 hucMSC의 지방형성 분화를 검출하였다. CD73, CD166(둘 다 1:10, BD Biosciences, Franklin Lakes, NJ, USA) 및 CD105(1:20, AbD Serotec, Oxford, 영국).
잘 자라는 hucMSC를 배양했습니다. 상층액을 수집하고 10cm 원심분리기 반경으로 1시간 동안 28,500r/min의 속도로 원심분리했습니다. 상층액은 버리고 세포를 2% glutaraldehyde와 1% osmic acid로 고정하고 에탄올로 탈수하고 propylene oxide에 담그고 2시간 동안 건조시킨 후 Epon812로 포매하여 얇게 썬다. 슬라이스는 각각 우라늄과 납으로 염색되었습니다. 마지막으로 전자현미경으로 엑소좀을 관찰하였다. Nanosight 검출기(Malvern Instruments, Malvern, UK)는 엑소좀 나노입자 및 그 크기의 브라운 운동 영상을 검출하는 데 사용되었습니다. hucMSC-Exos의 표면 마커는 western blot assay로 동정한 결과 hucMSC-Exos가 CD9, CD81, CD63을 발현하는 것으로 나타났다.
HucMSC는 miR-18b-3p 벡터의 낮은 발현과 miR-18b-3p 벡터 음성 대조군(NC)의 낮은 발현을 포함하는 렌티바이러스에 감염되었습니다(Shanghai GenePharma Co, Ltd, Shanghai, China). 마지막으로, 안정적으로 발현된 hucMSC-antagomir NC 및 hucMSC-miR-18b-3p antagomir를 얻었다. 세포를 48시간 동안 배양하고, 상등액을 수집하고 초원심분리로 원심분리하여 상응하는 Exos-antagomir NC 및 Exos-miR-18b-3p antagomir를 얻었다.
건강 상태가 깨끗하고 성적 성숙도가 높은 Wistar 쥐(무게 200–250g, 노화 8w, 성별)를 선택했습니다(중국 우한 소재 무한 대학 실험 동물 센터). 쥐는 18~28°C의 온도, 40~70%의 상대 습도, 적절한 식단과 물을 갖춘 차단 시스템에서 먹였습니다.
쥐의 PE 모델은 기사를 참조하여 50mg/kg의 산화질소 합성효소 억제제인 N(G)-nitro-l-arginine methyl ester(L-NAME, Beyotime)를 복강 내 주사하여 확립되었습니다[17]. PE 모델의 성공적인 확립은 20mmgHg 및 115mmHg 이상의 혈압 증가와 단백뇨 증가를 기반으로 했습니다.
암컷 쥐와 수컷 쥐를 1:1로 무작위 동거시켰고, 두 쥐는 오후 5~6시에 개별 특수 케이지에 보관했습니다. 전날. 암컷 쥐의 질분비물에 있는 정자는 다음날 질마개와 현미경으로 관찰하였다. 결과가 동시에 양성이면 그 날을 임신 0일로 기록하였다. 13 일 부터 임신일에 랫트를 6개 그룹(각 그룹당 10마리):정상 그룹(임신 13일부터 20일까지 동일한 양의 생리 식염수를 복강 내 주사), PE 그룹(L-NAME [50mg/ kg/day]는 임신 13일부터 20일까지 복강 주사하고, 임신 16일부터 19일까지는 생리식염수 20μL를 태반에 주사), PE + miR-NC군(L-NAME [50 mg /kg/day]를 임신 13일부터 20일까지 복강 주사하고, 4 nmol miR-NC를 임신 16일부터 19일까지 태반에 20 μL 주입), PE + miR-18b-3p agomir 군 (L-NAME[50mg/kg/day]은 임신 13일부터 20일까지 복강 내 주사하고, 20 μL의 4 nmol miR-18b-3p agomir를 임신 16일부터 19일까지 태반에 주사했습니다. , PE + miR-18b-3p 안타고미르군(L-NAME[50mg/kg/day]은 임신 13일부터 20일까지 복강 내 주사하고, 4 nmol miR-18b-3p 안타고미르 20 μL를 피 임신 16일 ~ 19일에 lacenta), PE + miR-18b-3p antagomir + small interfering RNA (si)-LEP 그룹(L-NAME [50mg/kg/day]은 13일부터 20일까지 복강 내 주사했습니다. 20 μL의 4 nmol miR-18b-3p antagomir 및 si-LEP를 임신 16일부터 19일까지 태반에 주사했습니다) 및 PE + si-LEP 그룹(L-NAME [50 mg/kg day]는 임신 13일부터 20일까지 복강 내 주사하고, 4 nmol si-LEP 20 μL를 임신 16일부터 19일까지 태반에 주사했습니다. 쥐는 렌티바이러스를 운반하는 엑소좀과 엑소좀으로 처리되었습니다. 랫트를 5개 그룹(각 그룹당 10마리):정상 그룹(임신 13일부터 20일까지 동일한 양의 생리 식염수를 복강 내 주사), PE 그룹(L-NAME(50mg/kg/day) )은 임신 13일부터 20일까지 복강 내 주사하고, 임신 16일부터 19일까지는 생리식염수 20μL를 태반에 주입), PE + Exos군(L-NAME(50mg/kg/day)) 임신 13일부터 20일까지 복강 내 주사하고, 20 μL의 Exos(80 μg 엑소좀은 20 μL 생리 식염수에 현탁)를 임신 16일부터 19일까지 태반에 주입했습니다. PE + Exos-antagomir NC 그룹(L-NAME(50mg/kg/day))은 임신 13일부터 20일까지 복강 내 주사하고, 태반에 Exos-antagomir NC(80μg 엑소좀을 생리식염수 20μL에 현탁) 20μL를 주입했습니다. 임신 16일부터 19일까지) 및 PE + Exos-miR-18b-3p 안타고미르 군(L-NAME(50mg/kg/일)을 13일부터 복강 내 주사 임신 20일까지, 그리고 20 μL의 Exos-miR-18b-3p antagomir(80 μg 엑소좀은 20 μL 생리 식염수에 현탁됨)를 임신 16일부터 19일까지 태반에 주입했습니다.
랫트의 압력은 랫트 꼬리동맥 혈압 측정으로 측정하였다. 임신 10, 13, 16 및 19일에 모든 임신한 쥐의 꼬리 커프 SBP를 쥐 꼬리 동맥 압력 검출기(Tensys (R) Medical Inc., San Diego, CA, USA)를 사용하여 측정했습니다. 압력은 짧은 시간에 3번 측정되었습니다. 그런 다음 평균값을 혈압으로 취했습니다.
무상 식이와 물의 경우 임신 10일, 13일, 16일, 19일에 임신한 쥐의 24시간 소변을 채취해 우한대학교 인민병원 신장과에서 단백질 함량을 검출했다.
임신 21일째에 임신한 쥐를 3% 펜토바르비탈 나트륨으로 마취시켰다. 쥐의 말초혈액은 보존하고 원심분리하여 혈청을 채취하여 -20°C의 냉장고에 보관하여 대기하였다. 그런 다음 제왕절개를 통해 랫트 태아와 태반을 채취하고 태아막과 연결된 탯줄을 제거하고 랫트 태아에 연결된 탯줄을 절단하였다. 태반과 태아 래트를 각각 무균 거즈에 올려 혈액과 양수를 건조시킨 후, 분석 저울에 올려 무게를 쟀다. 태반 조직의 한 부분을 4% 파라포름알데히드로 고정하고, 에탄올로 탈수하고, 자일렌으로 제거하고, 파라핀으로 포매하고, 헤마톡실린-에오신(HE) 염색 및 말단 데옥시뉴클레오티딜 트랜스퍼라제-매개 데옥시우리딘 트리포스페이트-비오틴을 위해 연속적으로 교차 슬라이스(5μm) nick end-labeling(TUNEL) 염색. 나머지는 역전사 정량적 중합효소 연쇄 반응(RT-qPCR) 검출, 웨스턴 블롯 분석 및 효소 결합 면역흡착 분석(ELISA)을 위해 -80°C에 보관되었습니다.
종양 괴사 인자-α(TNF-α), 인터루킨(IL)-1β 및 혈청 내 IL-6 함량을 ELISA로 테스트했습니다. TNF-α, IL-1β 및 IL-6의 농도는 키트(R&D Systems, Minneapolis, MN, USA)의 지침에 따라 결정되었습니다. 광학 밀도(OD) 값(490nm)은 마이크로플레이트 판독기(Thermo Fisher Scientific, MA, USA)로 테스트했습니다. OD 값을 횡축으로 하고 해당 표준 시료의 농도를 종축으로 하여 해당 표준 곡선을 구했습니다. TNF-α, IL-1β 및 IL-6 농도는 표준 곡선에서 계산되었습니다.
태반 조직의 파라핀 샘플은 자일렌으로 제거하고, 기존의 구배 알코올로 탈수하고, 헤마톡실린으로 염색하고, 1% 염산 알코올로 구별하고, 1% 암모니아수에 의해 파란색으로 되돌렸습니다. 그런 다음 조직을 1% 에오신 용액으로 대조염색하고 탈수(각각 75%, 90%, 95% 에탄올, 무수 에틸 알코올)하고 자일렌으로 제거하고 건조하고 차단하고 전자 현미경으로 관찰했습니다.
파라핀이 포매된 절편을 지침에 따라 일상적으로 탈랍 및 탈수한 다음 TUNEL Kit(Nanjing Kejin Biotechnology Co., Ltd., Jiangsu, China)로 세포자멸사를 감지했습니다. 4,6-Diamino-2-phenylindole(Shanghai Baitai Biotechnology Co., Ltd., Shanghai, China)는 형광 현미경(Nikon, Tokyo, Japan)을 사용하여 TUNEL 양성 세포를 관찰하는 데 사용되었습니다[18].
태반 조직의 무게를 측정했습니다. 50~100mg의 태반 조직에 1mL의 TRIzol(Invitrogen, Carlsbad, California, USA)을 첨가하고 완전히 용해했습니다. 조직에 200μL 클로로포름을 첨가하고 4°C, 12,000rpm에서 원심분리하여 총 RNA를 추출했습니다. RNA의 농도 및 순도는 DU-800 단백질 핵산 분광광도계(Beckman)로 측정하였다. U6 및 β-액틴을 로딩 대조군으로 사용하였다. PCR 프라이머는 Shanghai Sangon Biotechnology Co. Ltd.(중국 상하이)에서 설계하고 합성했습니다. 프라이머 서열은 표 1에 나열되어 있습니다. RNA는 RNA 역전사 키트(Sangon)의 지침에 따라 cDNA로 역전되었습니다. PCR을 증폭하고 생성물을 아가로스 겔 전기영동으로 확인하였다. 데이터는 2 −ΔΔCt 로 계산되었습니다. 방법.
태반 조직의 총 단백질은 방사성 면역 침전 분석 세포 용해 완충액(Beyotime)에 의해 추출되었습니다. HucMSC-Exo는 14,000rpm에서 원심분리된 버퍼를 추출하는 데 사용되었습니다. 혈청에서 엑소좀 마커 단백질(CD81, CD63 및 CD9)의 단백질 발현을 테스트하기 위해 상청액을 보존했습니다. 단백질 농도는 비신코닌산 키트(Beyotime, P0010)로 측정하였다. 단백질의 정량적 결과에 따라 시료를 로딩하고 10% sodium dodecyl sulfate-polyacrylamide gel 전기영동으로 처리하여 멤브레인으로 옮겼다. 막을 5% 탈지유로 차단하고 1차 항체 LEP, CD63, CD81, CD9 및 β-액틴(4mL, 1:1000, Santa Cruz Biotechnology, Inc, Santa Cruz, CA, USA)으로 프로브하고 다시 프로브했습니다. 4mL 2차 항체 염소 항토끼 IgG/양고추냉이 퍼옥시다제를 사용하여 노출 및 개발했습니다. β-액틴은 내부 참조로 활용되었습니다. 회색 값은 겔 그래픽 분석 소프트웨어 Image Lab으로 분석되었습니다.
온라인 예측 소프트웨어 https://cm.jefferson.edu/를 채택하여 miR-18b-3p와 LEP 사이의 표적 관계 및 miR-18b-3p와 LEP 3' untranslated region(UTR)의 결합 부위를 예측했습니다. ). miR-18b-3p 결합 부위를 포함하는 LEP 3'UTR 프로모터 영역 서열을 구성하였다. LEP 3'UTR 야생형(WT) 플라스미드 및 LEP 3'UTR 돌연변이 유형(MUT)을 구축했습니다. 재조합 플라스미드는 각각 LEP 3'UTR-WT 및 LEP 3'UTR-MUT로 명명되었습니다. 배양된 293T 세포를 miR-18b-3p 모방체 및 LEP 3'UTR-WT, miR-18b-3p 모방체 및 LEP 3'UTR-MUT, 모방 NC 및 LEP 3'UTR-WT, 모방 NC 및 30시간 동안 LEP 3'UTR-MUT. 그런 다음 293T 세포를 수집했습니다. 세포에서 반딧불이 및 레닐라 루시퍼라제 활성은 이중 루시페라제 리포터 유전자 검출 키트(Promega, Madison, WI, USA)에 따라 발광 측정에 의해 측정되었습니다.
Biotinylated RNA 프로브(Bio-miR-NC, Bio-miR-18b-3p 및 Bio-miR-18b-3p-Mut)를 293T 세포의 용해물과 함께 배양하고 항생제 스트렙토마이신이 접합된 자기 비드를 사용하여 추출했습니다. 실험은 Pierce 자기 RNA 풀다운 키트(Pierce, IL, USA)의 지침에 따라 수행되었습니다. RNA는 TRIzol(Pierce)을 사용하여 용출 및 정제되었습니다. RNA 복합체에서 LEP의 농축은 이전에 설명한 대로 RT-qPCR을 사용하여 정량화되었습니다[19].
모든 데이터는 SPSS 21.0 소프트웨어(IBM Corp. Armonk, NY, USA)에 의해 설명되었습니다. 측정 데이터는 평균 ± 표준편차로 표시하였다. 데이터는 독립 표본 t에 의해 수행되었습니다. 두 그룹 비교를 위한 테스트인 반면, 여러 그룹 간의 비교는 일원 분산 분석(ANOVA)에 이어 Tukey의 사후 테스트로 평가되었습니다. 통계적 유의성에 대한 기준은 p로 설정되었습니다. <0.05.
<섹션 데이터-제목="결과">도립현미경으로 탯줄 조직 덩어리를 관찰하였다. 3일째에 세포가 조직 덩어리에서 기어나오는 것을 볼 수 있었습니다. 세포는 약 5일째에 방추형과 실 모양을 보였을 뿐만 아니라 콜로니처럼 성장하였다. 3번 계대까지 배양하였을 때 세포의 형태는 균일한 긴 방추형이었고 섬유아세포 형태와 유사하였고 배열은 규칙적이었다(Fig. 1a). hucMSC의 지방 분화 2w 후, 세포질에 지질 방울이 형성되었고, 지질 방울은 도립 현미경(그림 1b)에서 Kranz 구조를 보여, 분리된 배양 hucMSC가 지방 분화 능력이 있음을 시사합니다. 2w의 골형성 분화 후, 다수의 갈색 칼슘 결절이 도립현미경 하에서 관찰될 수 있었고(도 1c), 분리된 배양 hucMSC가 골형성 분화 능력을 가짐을 나타낸다. 세포의 면역 표현형을 테스트하기 위해 유세포 분석기를 채택했으며 그 결과 세포가 MSC의 표면 마커 CD73, CD105 및 CD166을 과발현하는 것으로 나타났습니다(그림 1d).
<그림>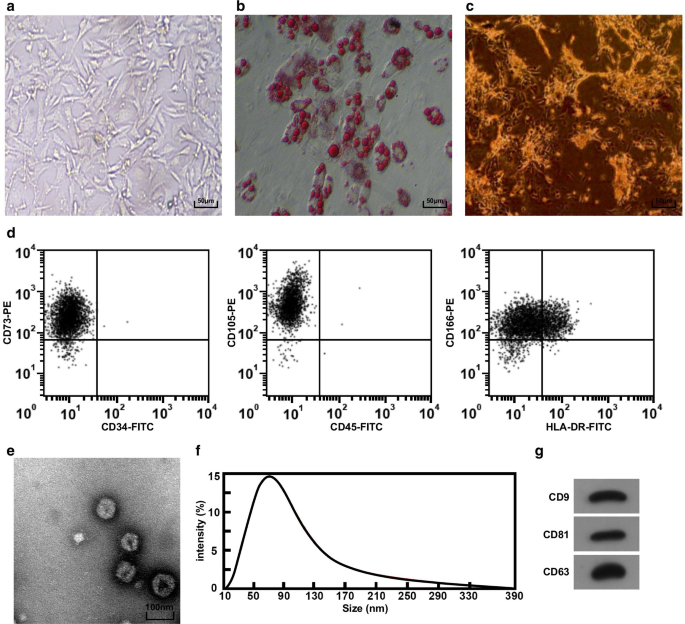
hucMSC 및 hucMSC-Exos의 형태 및 식별. 아 도립 현미경에서 hucMSC의 형태, b hucMSC는 오일 레드 O 염색으로 테스트되었습니다. ㄷ hucMSC는 ALP 염색으로 테스트되었습니다. d 유동 세포 계측법을 사용하여 면역 표현형을 검출했습니다. 이 TEM으로 관찰한 hucMSC-Exos의 모양과 크기. 에 나노사이트 분석을 이용한 엑소좀 입자 크기 분포 검출 지 hucMSC-Exos에서 CD9, CD81 및 CD63의 단백질 발현은 웨스턴 블롯 분석으로 검출되었습니다.
hucMSC-Exos의 형태는 TEM에 의해 관찰되었으며, 그 결과 엑소좀은 중앙 밀도가 낮고 양쪽에 두꺼운 염색이 있는 원형 또는 타원형임을 나타냈다(그림 1e). 나노사이트 분석을 이용하여 엑소좀의 입자 크기를 분석한 결과, 입자 크기가 주로 40~100nm 사이에 분포하고 80nm 부근에 더 집중된 것으로 나타났습니다(그림 1f). Western blot assay에서는 모든 표면 마커인 CD81, CD63, CD9가 hucMSC-Exos에서 발현되는 것으로 나타났습니다(그림 1g).
SBP 및 24시간 단백뇨의 결과는 투여 전(임신 10일째) 6군에서 SBP 및 24시간 단백뇨에 유의한 차이가 없었다는 것을 나타냈다. 임신 19일째의 SBP와 24시간 단백뇨는 정상 쥐에서 뚜렷한 차이를 보이지 않았습니다. miR-NC, miR-18b-3p agomir, miR-18b-3p antagomir, miR-18b-3p antagomir + si-LEP, si-NC 또는 si-LEP, SBP 및 24-h로 처리된 PE 래트 또는 PE 래트에서 단백뇨는 임신 13일째에 증가하기 시작했습니다. miR-18b-3p agomir와 si-LEP로 처리된 PE 쥐에서 임신 16일과 19일에 SBP와 24시간 단백뇨의 뚜렷한 차이는 없었습니다. PE 쥐는 임신 19일에 SBP와 24시간 단백뇨가 증가했습니다. 이 증가는 miR-18b-3p 상승에 의해 감소되었지만 miR-18b-3p 억제에 의해 더욱 강화되었습니다. LEP 감소는 PE 쥐에서 임신 19일째에 SBP 및 24시간 단백뇨에서 miR-18b-3p 하향 조절의 역할을 폐지했습니다(그림 2a, b).
<사진>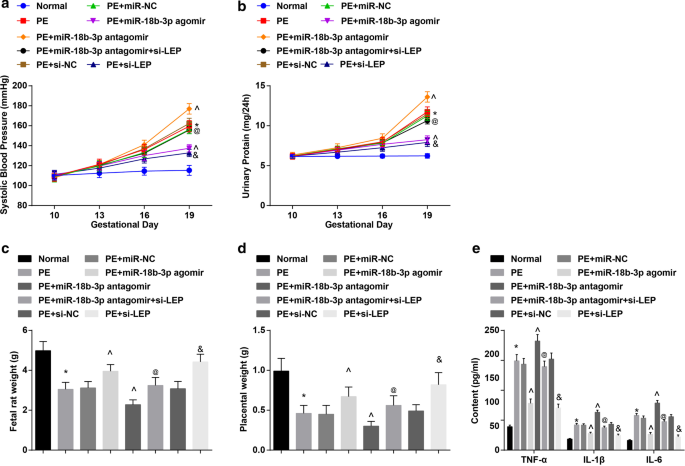
miR-18b-3p를 복원하면 PE 쥐의 병리학적 특성이 완화됩니다. 아 쥐의 SBP 결과. ㄴ 쥐의 24시간 단백뇨 결과. ㄷ 태아 쥐의 체중 변화. d 쥐의 태반 무게 변화. 이 혈청 내 염증 인자의 변화를 ELISA를 이용하여 검출하였다. n =10, *p <0.05 대 정상 그룹. ^p <0.05 대 PE + miR-NC 그룹. @ 피 <0.05 대 PE + miR-18b-3p 안타고미르 그룹. & 피 <0.05 대 PE + si-NC 그룹. 측정 데이터는 평균 ± 표준편차로 표시하였으며, 여러 그룹 간의 비교는 일원 분산분석(one-way ANOVA) 후 Tukey의 검정으로 평가했습니다.
PE 쥐에서 태아 쥐와 태반의 무게가 감소했습니다. 상향 조절된 miR-18b-3p 또는 하향 조절된 LEP는 증가하는 반면 하향 조절된 miR-18b-3p는 PE 쥐에서 태아 쥐와 태반의 무게를 감소시켰습니다. LEP 침묵은 PE 쥐의 태아 쥐와 태반의 무게에 대한 miR-18b-3p 억제 효과를 역전시켰습니다(그림 2c, d).
PE 쥐의 혈청에서 염증 인자가 검출되었습니다. PE 래트에서 TNF-α, IL-1β 및 IL-6 함량이 향상되었음을 발견했습니다. miR-18b-3p 증가 또는 LEP 억제는 억제된 반면 miR-18b-3p 감소는 TNF-α, IL-1β 및 IL-6의 함량을 촉진했습니다. TNF-α, IL-1β 및 IL-6 함량에 대한 억제된 miR-18b-3p의 효과는 LEP 고갈에 의해 폐지되었습니다(그림 2e).
정상 쥐에서 태반 융모는 혈관이 풍부하고 명확한 구조를 가졌고, 합포체영양막은 태반 융모의 주요 영양막이었고, 세포영양막은 더 적었다. miR-NC, miR-18b-3p antagomir, si-NC 또는 miR-18b-3p antagomir + si-LEP로 처리된 PE 랫트 또는 PE 랫트에서 태반 융모의 수가 감소하고 구조가 흐려지고 위축되었으며 일부 융모 섬유소성 괴사를 시행하였고 태반 융모의 융합체영양막 결절의 수가 증가하였고 융모의 대부분이 미성숙하였다. miR-18b-3p agomir와 si-LEP를 처리한 PE 쥐에서 영양 세포의 수가 감소하고 병리학적 변화가 완화되었습니다(그림 3a).
<그림>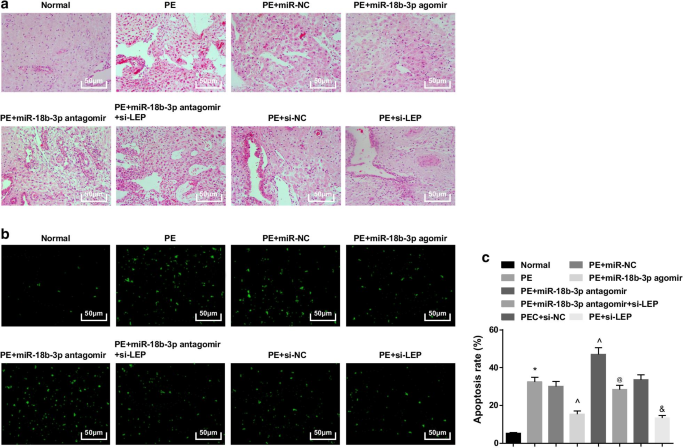
과발현된 miR-18b-3p는 병리학적 변화를 개선하고 PE 쥐에서 태반 조직의 세포 사멸 세포를 억제합니다. 아 H 염색은 태반 조직의 병리학적 특징을 테스트하는 데 사용되었습니다. ㄴ TUNEL 염색은 PE 쥐에서 태반 조직의 세포 사멸 세포를 결정하기 위해 구현되었습니다. ㄷ TUNEL 염색을 통해 세포의 세포자살률을 확인하였다. n =10, *p <0.05 대 정상 그룹. ^p <0.05 대 miR-NC 그룹. @ 피 <0.05 대 miR-18b-3p antagomir 그룹. & 피 <0.05 대 si-NC 그룹. 측정 데이터는 평균 ± 표준편차로 표시하였으며, 여러 그룹 간의 비교는 일원 분산분석(one-way ANOVA) 후 Tukey의 검정으로 평가했습니다.
TUNEL 염색은 소수의 사멸 세포가 보일 수 있음을 시사했습니다. PE 쥐는 miR-18b-3p 상승과 LEP 침묵에 의해 감소된 세포 사멸 세포가 증가했으며 miR-18b-3p 억제에 의해 더욱 강화되었습니다. LEP 침묵은 또한 PE 쥐의 세포 사멸 세포 수에 대한 miR-18b-3p 억제 효과를 역전시켰습니다(그림 3b, c).
종합하면, miR-18b-3p가 상향 조절되거나 LEP가 억제된 쥐는 조직학에서 PE 진행 정도가 감소했으며, 침묵된 LEP는 억제된 miR-18b-3p의 치료 효과를 없앨 수 있었습니다.
위의 결과에 기초하여 LEP 하향 조절은 병리학 및 조직학에서 PE 래트에 대한 miR-18b-3p 하향 조절의 치료 효과를 역전시켰습니다. 따라서 우리는 miR-18b-3p가 LEP와 관련이 있을 수 있다고 가정했습니다.
Western blot assay와 RT-qPCR은 PE 쥐가 miR-18b-3p를 감소시키고 LEP 발현 수준을 증가시킨 것으로 나타났습니다. miR-18b-3p agomir의 치료는 PE 쥐에서 miR-18b-3p를 상향 조절하고 LEP를 하향 조절한 반면 miR-18b-3p antagomir의 치료는 LEP 발현을 증가시켰습니다. LEP 침묵은 PE 쥐의 LEP 발현에 대한 miR-18b-3p 감소의 촉진 효과를 역전시켰습니다(그림 4a-c).
<그림>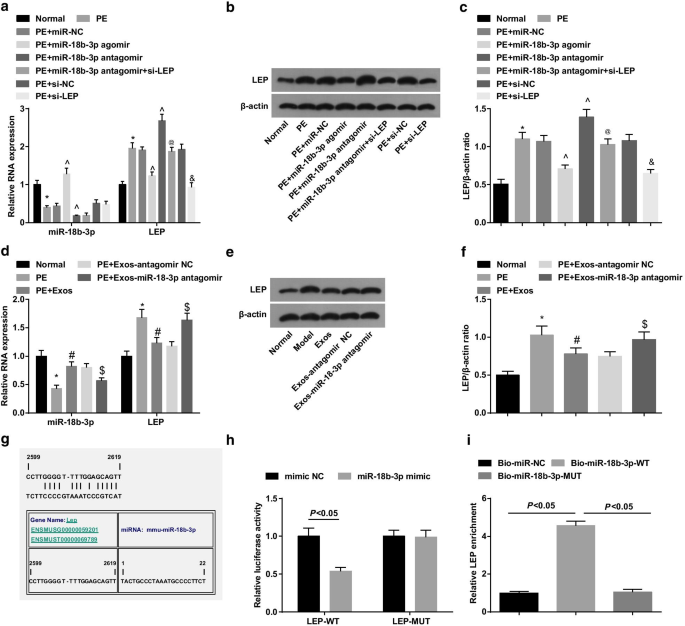
miR-18b-3p는 하향 조절되고 LEP는 PE 쥐의 태반 조직에서 상향 조절됩니다. 아 태반 조직에서 miR-18b-3p 및 LEP mRNA의 발현은 RT-qPCR을 사용하여 검출되었습니다. ㄴ 태반 조직에서 LEP의 단백질 밴드. ㄷ 태반 조직에서 LEP 단백질 발현을 검출하기 위해 웨스턴 블롯 분석을 수행하였다. d 엑소좀 처리 후 태반 조직에서 miR-18b-3p 및 LEP mRNA의 발현을 RT-qPCR을 사용하여 검출하였다. 이 엑소좀 처리 후 태반 조직에서 LEP의 단백질 밴드. 에 엑소좀 처리 후 LEP 단백질 발현을 검출하기 위해 웨스턴 블롯 분석을 수행하였다. 지 온라인 소프트웨어에 의해 예측되는 miR-18b-3p 및 LEP의 결합 부위. 아 이중 루시퍼라제 리포터 유전자 분석으로 검증된 miR-18b-3p와 LEP 간의 표적 관계. 나 miR-18b-3p와 LEP 사이의 표적화 관계는 RNA 풀다운 분석으로 확인되었습니다. n =10, *p <0.05 대 정상 그룹. ^p <0.05 대 miR-NC 그룹. @ 피 <0.05 대 miR-18b-3p antagomir 그룹. & 피 <0.05 대 si-NC 그룹. # 피 <0.05 대 PE 그룹. $ 피 <0.05 대 PE + Exos-antagomir NC 그룹. 측정 데이터는 평균 ± 표준편차로 표시하였으며, 여러 그룹 간의 비교는 일원 분산분석(one-way ANOVA) 후 Tukey의 검정으로 평가했습니다.
웨스턴 블롯 분석 및 RT-qPCR을 사용하여 PE 래트에서 엑소좀의 역할을 조사했습니다. 결과는 엑소좀이 PE 쥐에서 miR-18b-3p를 상향 조절하고 LEP를 하향 조절하여 PE 발달에 대한 엑소좀의 억제 효과를 나타내는 것으로 나타났습니다. 또한 miR-18b-3p antagomir를 전달하는 엑소좀은 PE 쥐에서 miR-18b-3p 하향 조절 및 LEP 상향 조절을 유도했습니다(그림 4d-f).
miR-18b-3p와 LEP 사이의 목표 관계는 생물정보학 온라인 예측 소프트웨어 https://cm.jefferson.edu/에 의해 예측되었습니다(그림 4g). 이중 루시퍼라제 리포터 유전자 분석은 miR-18b-3p 모방이 LEP 3'UTR-WT의 루시퍼라제 활성을 감소시키는 반면 LEP 3'UTR-MUT의 루시퍼라제 활성에는 영향을 미치지 않는다고 제안했습니다(그림 4h). 또한, RNA 풀다운 분석은 LEP 농축이 WT-비오틴화된 miR-18b-3p에 의해 증가되었음을 보여주었습니다(그림 4i). 이러한 결과는 LEP가 miR-18b-3p의 표적 유전자임을 나타냅니다.
SBP 및 24시간의 결과는 투여 전(임신 10일째) 5군에서 SBP 및 24시간 단백뇨에 유의한 차이가 없었다는 것을 제시하였다. SBP and 24-h proteinuria in day 19 of gestation showed no distinct difference in normal rats. In PE rats, SBP and 24-h proteinuria began to raise at day 13 of gestation. There was no distinct difference of SBP and 24-h proteinuria in day 16 and day 19 of gestation in PE rats treated with hucMSC-Exos and hucMSC-Exos transmitting antagomir NC. SBP and 24-h proteinuria heightened in day 19 of gestation in the PE rats, while the increase was reduced by injection of hucMSC-Exos. Inhibiting miR-18b-3p reversed the effect of hucMSC-Exos on SBP and 24-h proteinuria in day 19 of gestation in PE rats (Fig. 5a, b).

hucMSC-Exos attenuate pathological characteristics of PE rats. 아 Results of SBP in rats after exosome treatment. ㄴ Results of 24-h proteinuria in rats after exosome treatment. ㄷ Weight changes of fetal rats after exosome treatment. d Changes of placental weight in rats after exosome treatment. 이 Changes of inflammation factors after exosome treatment in serum were determined using ELISA. n = 10, *p < 0.05 versus the normal group. # 피 < 0.05 versus the PE group. $ 피 < 0.05 versus the PE + Exos-antagomir NC group. Measurement data were depicted as mean ± standard deviation, and comparisons among multiple groups were assessed by one-way ANOVA followed by Tukey’s test
The weight of fetal rat and placenta was measured, and we found that the PE rats had decreased weight of fetal rat and placenta; miR-18b-3p downregulation abolished the role of hucMSC-Exos in the weight of fetal rat and placenta in PE rats (Fig. 5c, d).
Inflammatory factors in serum were detected using ELISA. TNF-α, IL-1β and IL-6 contents remarkably increased in PE rats. Exosomes treatment decreased TNF-α, IL-1β and IL-6 contents in serum of PE rats, which were enhanced by injection of exosomes inhibiting miR-18b-3p (Fig. 5e).
In normal rats, the placental villus was abundant in blood vessels with a clear structure, syncytiotrophoblasts were the main trophoblast in placental villi, and there were fewer cytotrophoblasts. In the PE rats and PE rats treated with hucMSC-Exos-miR-18b-3p-antagomir, the number of placental villi reduced, the structure was blurred and atrophied, some villi were presented fibrinoid necrosis, and the number of syncytiotrophoblast nodules in placental villi enhanced, and most of the villi were immature. The pathological change was improved in the PE rats treated with hucMSC-Exos or hucMSC-Exos-antagomir NC versus the PE rats and PE rats treated with hucMSC-Exos-miR-18b-3p antagomir (Fig. 6a).

Exosomes alleviate pathological change and decrease apoptotic cells of placenta tissues in PE rats. 아 HE staining was utilized to test pathological features of placenta tissues in PE rats after exosome treatment. ㄴ TUNEL staining was implemented to determine apoptotic cells of placenta tissues in PE rats after exosome treatment. ㄷ Cell apoptosis rate was detected by TUNEL staining. n = 10, *p < 0.05 versus the normal group. # 피 < 0.05 versus the PE group. $ 피 < 0.05 versus the PE + Exos-antagomir NC group. Measurement data were depicted as mean ± standard deviation, and comparisons among multiple groups were assessed by one-way ANOVA followed by Tukey’s test
TUNEL staining indicated that in normal rats, a small number of apoptotic cells could be seen. PE rats had enhanced apoptotic cells, and reduced miR-18b-3p reversed the impacts of hucMSC-Exos on the number of apoptotic cells in placenta tissues from PE rats (Fig. 6b, c).
PE is a multisystem pregnancy disorder characterized by proteinuria and either high blood pressure or other adverse conditions and is linked to a wide range of maternal endothelial dysfunction [20]. It was reported that hucMSC-Exo improved the morphology of placental tissue in PE rats through suppressing cell apoptosis and facilitating angiogenesis in placental tissue in a dose-dependent manner [8]. A study has reported that miR-18b expression affected cell migration, viability and invasion in PE [12]. Moreover, it was verified increased maternal LEP concentration and hypomethylation of the LEP in placenta in early onset PE [21]. The current study was designed to explore the effect of exosomes and miR-18b-3p targeted LEP on the occurrence of PE. The findings in this study revealed that hucMSC-derived exosomal miR-18b-3p inhibited PE progression by reducing LEP.
Based on our findings, miR-18b-3p reduced and LEP elevated in placenta tissues of PE rats. Similar to our study, the mRNA expression of miR-18b was markedly suppressed in PE placental tissues relative to that in normal placental tissues [12]. In addition, a study revealed that miR-18b content was dramatically reduced in malignant melanoma tissues in comparison with their matched adjacent non-tumor tissues [22]. Another study has verified that placental LEP expression was raised in preterm PE compared with controls [23]. Moreover, a study showed that LEP expression was obviously heightened in preeclamptic placentas [15]. This literature provided a theoretical basis for us to explore the abnormal expression of miR-18b-3p and LEP in PE. Moreover, it was predicted using a bioinformatic software that LEP was targeted by miR-18b-3p, and this targeting relationship was further confirmed with dual-luciferase reporter gene assay in our research. A study reported that LEP is a target for all three miRNAs (miR-1301, miR-223 and miR-224) in early-onset PE [16]. Another study has displayed that LEP decreased miR-93 expression in osteoarthritis and rheumatoid arthritis [24]. However, the binding between miR-18b-3p and LEP in human diseases, especially in PE, remains scarcely studied, which is the novelty of this study. Furthermore, a result emerging from our study reported that exosomes increased miR-18b-3p and decreased LEP in placenta tissues of PE. It was formerly documented that the expression of miR-18b-5p was notably raised in colorectal cancer plasma exosomes [25], while the relationship between hucMSC-Exos and miR-18b-3p/LEP in PE needs further study.
Additionally, the finding from our investigation showed that restored miR-18b-3p reduced SBP and 24-h proteinuria of PE rats, increased the weight of placenta, declined TNF-α, IL-1β and IL-6 contents in serum and placenta tissues as well as suppressed cell apoptosis. These data indicated that miR-18b-3p elevation contributes to alleviating the symptoms and pathological changes in PE. It was demonstrated that stable upregulation of miR-18b produced effective tumor inhibitor activity, such as inhibiting melanoma cell viability, inducing apoptosis and reducing tumor growth in vivo [26]. Another result in our study was that hucMSC-Exos reduced SBP and 24-h proteinuria of PE rats, increased the weight of placenta, declined TNF-α, IL-1β and IL-6 contents in serum and placenta tissues as well as suppressed cell apoptosis. The findings of the current study revealed that exosomes treated PE rat models presented an increase of the number and quality of fetuses, the quality of placenta, but cell apoptosis was significantly reduced [8]. Interestingly, a previous research has demonstrated that the addition of fetal bovine exosomes declined contents of macrophage TNF-α and IL-6 [27]. A study has revealed that purified exosomes suppressed production of IL-1β in lipopolysaccharide/nigericin-stimulated macrophages [28]. Furthermore, Nong et al. have suggested that inflammatory markers, such as TNF-α and IL-6, were dramatically decreased after administration of exosomes produced through human-induced pluripotent stem cell-derived MSCs [29]. There is a article finding that the SBP was markedly elevated in the group of women who later developed PE [30, 31]. It was displayed that PE patients were positively associated with SBP and diastolic blood pressures and proteinuria [32]. Also, a recent study has provided a proof that proteinuria heightened with advancing gestation in PE women [33]. A important finding was that rats from the PE group had increased TNF-α relative to the normal pregnant group [34]. Another study has verified that serum IL-6 and IL-1β were obviously elevated in women with PE in relation to controls [35]. The above findings suggested that PE patients usually showed high SBP, proteinuria and levels of inflammatory factors. Thus, it could be inferred from our results that the hucMSC-derived exosomal miR-18b-3p had a therapeutic effect on PE.
In conclusion, our study provides evidence that hucMSCs-derived exosomes upregulate miR-18b-3p, which targets LEP to suppress the contents of inflammatory factors and reduce cell apoptosis rate in PE rat placenta tissues, thereby inhibiting the occurrence of PE. Thus, exosomal miR-18b-3p may be a potential candidate for treatment of PE via targeting LEP. This research identified the role of hucMSC-derived exosomal miR-18b-3p targeting LEP during PE development for the first time, which provided a novel insight for PE treatment. However, due to the limitation of known researches, the study needs to be monitored rigorously and reported appropriately in the future clinical trials.
Not applicable.
나노물질
RPA 고객 서비스, 로봇 호출 고객 서비스와 디지털 혁신 – 오늘날 어떤 기업도 무시할 수 없는 두 가지 용어입니다. 비즈니스를 운영한다는 모든 아이디어는 고객을 중심으로 이루어집니다. 그리고 수익성을 달성하려는 목표는 현대 기술을 올바르게 사용하여 달성할 수 있습니다. 이 두 가지를 분리하는 것은 불가피합니다. 사실, 그것들을 하나로 모으면 놀라운 일이 일어날 수 있습니다. 로봇 프로세스 자동화란 무엇입니까? 이 게시물에서 매우 많이 찾는 기술 중 하나를 로봇 프로세스 자동화 또는 RPA라고 합니다. IEEE Working
기업은 인적 자본으로 대표되는 부를 무시할 수 없습니다. 꿈꾸고, 직관하고, 위험을 감수하고, 무엇보다도 언제 재앙을 향해 가고 있는지 인식하는 것 지난 여름은 인공 지능 역사에서 암울한 이정표를 보았습니다. 자율 주행 차량으로 인한 첫 번째 사망이었습니다. 완전 자동 조종 장치로 작동하는 Tesla Model S가 플로리다 고속도로에서 트럭 뒤쪽에 충돌하여 인간 운전자가 사망했습니다. 회사 블로그에 따르면 이 비극적인 사고의 원인은 AI가 밝은 하늘을 배경으로 트랙터 트레일러의 흰색 면을 찾아내지 못했기 때문입니다. Tesla