억제된 MicroRNA-301은 PTEN을 증가시켜 식도 편평 세포 암종의 혈관 신생 및 세포 성장을 억제합니다.
초록
목표
식도 편평 세포 암종(ESCC)은 조기 전이와 후기 진단이 특징입니다. MicroRNA-301(miR-301)은 다양한 암에 관여하는 것으로 알려져 있습니다. 그럼에도 불구하고 ESCC에 대한 miR-301의 영향은 아직 밝혀지지 않았습니다. 따라서 우리는 ESCC 진행에서 miR-301의 역할을 탐구하는 것을 목표로 합니다.
방법
ESCC 조직 및 세포주에서 miR-301 및 포스파타제 및 텐신 동족체(PTEN)의 발현을 평가하였다. 다음으로, 스크리닝된 세포에 변형된 miR-301 또는 PTEN 올리고뉴클레오티드 및 플라스미드를 처리한 후, ESCC 세포의 콜로니 형성 능력, 세포 생존력, 이동, 침입, 세포 주기 분포 및 세포자멸사를 평가하였다. 또한, 종양 성장 및 미세혈관 밀도(MVD)도 평가되었으며, miR-301과 PTEN 간의 표적화 관계가 확인되었습니다.
결과
MiR-301은 상향 조절되었고 PTEN은 ESCC 조직과 세포에서 하향 조절되었습니다. 기능 분석을 위해 KYSE30 세포 및 Eca109 세포를 선택했습니다. KYSE30 세포에서 억제된 miR-301 또는 과발현된 PTEN은 세포 악성 행동을 억제하고 침묵된 PTEN은 ESCC 진행에 대한 miR-301 억제의 영향을 제거했습니다. Eca109 세포에서 miR-301 과발현 또는 PTEN 억제는 세포 악성 행동을 촉진했으며 PTEN 과발현은 ESCC 진행에 대한 miR-301 상승 효과를 역전시켰습니다. 생체 내 분석은 miR-301 억제 또는 PTEN 과발현이 ESCC 종양 성장 및 MVD를 억제하고 miR-301 상승 또는 PTEN 감소가 반대 효과를 갖는 것으로 밝혀졌다. 게다가 PTEN은 miR-301의 표적이 되었습니다.
결론
종합하면, 우리 연구의 결과는 miR-301이 ESCC에서 PTEN 발현을 조절함으로써 세포 성장, 전이 및 혈관신생에 영향을 미치는 것으로 나타났습니다.
소개
식도암(EC)은 전 세계적으로 8번째로 흔한 암으로 사망률이 높고 예후가 좋지 않은 치명적인 악성종양이다[1]. 전체 EC 사례의 약 90%를 차지하는 식도 편평 세포 암종(ESCC)은 중국에서 EC의 주요 형태입니다[2]. 낮은 사회경제적 지위, 흡연, 음주, 열악한 영양섭취, 니트로사민이 풍부한 식품 또는 진균독소에 오염된 식품 등 다양한 원인이 ESCC를 유발한다[3]. ESCC 환자의 임상 결과와 투여가 촉진되었음에도 불구하고 5년 생존율이 15-25%로 여전히 예후가 좋지 않습니다[4]. 따라서 ESCC 환자에게 보다 효과적인 치료 방법을 제공하기 위해서는 ESCC 개발에서 바이오마커로 기능할 수 있는 종양 유전자 또는 종양 억제 유전자를 확인하는 것이 중요합니다.
MicroRNA(miRNA)는 유전자 발현 조절에 필수적인 역할을 하는 작은 비암호화 RNA이며[5], mRNA의 안정성과 mRNA의 능력을 조절하여 종양 진행에 영향을 미치는 능력이 있는 것으로 밝혀졌습니다[6]. miR-4324[7], miR-889-3p[8] 및 miR-9[9]와 같은 miRNA의 양이 ESCC의 과정과 관련이 있는 것으로 밝혀졌습니다. MiR-301은 인간 게놈의 17q22-23에 위치한 fam33a 전사 단위에 의해 형성되는 miRNA의 구성원입니다. miR-301의 과발현은 이전에 확인되었으며 이는 인간 질병과 관련이 있음을 반영합니다[6, 10]. 그러나 miR-301의 기능 메커니즘은 ESCC에서 밝혀지지 않았습니다. 또한, PTEN(phosphatase and tensin homologue)은 종양에서 자주 파괴되고 암 환자의 생식선 돌연변이에 의해 표적화되는 것으로 확인되어 종양의 억제 역할을 한다[11]. PTEN의 조절장애가 ESCC 발달과 상관관계가 있음이 검증되었습니다[12]. 흥미롭게도 최근 연구에서는 miR-301이 비소세포폐암에서 PTEN을 표적으로 한다는 사실이 밝혀졌습니다[13]. 그러나 ESCC 개발에서 miR-301과 PTEN 간의 이러한 표적화 관계는 아직 밝혀지지 않았습니다. 우리의 연구는 ESCC 진행에 대한 miR-301 및 PTEN의 영향에 초점을 맞추었으며, 이는 크게 알려지지 않고 참신합니다. 우리는 miR-301이 PTEN 발현을 조절함으로써 ESCC에서 혈관신생 및 세포 성장에 영향을 미칠 수 있다고 추론했습니다.
자료 및 방법
윤리 성명서
연구에 앞서 모든 환자로부터 서면 동의를 얻었습니다. 이 연구의 프로토콜은 길림대학교 제2병원 윤리위원회의 승인을 받았으며 헬싱키 선언의 인간 피험자를 대상으로 하는 의학 연구에 대한 윤리 원칙을 기반으로 합니다. 동물 실험은 국립 보건원에서 발행한 실험 동물의 관리 및 사용에 대한 지침과 엄격하게 일치했습니다. 동물 실험 프로토콜은 길림대학교 제2병원 동물관리위원회의 승인을 받았습니다.
연구 주제
1월부터 길림대학교 제2병원 흉부외과에서 식도절제술을 받은 ESCC 환자(남성 78명, 여성 32명)로부터 ESCC 조직 및 인접 정상 조직(종양으로부터> 5 cm) 샘플 100개를 수집했습니다. 2015년 ~ 2017년 12월. 110명의 환자 중 84건> 60세, 26건 ≤ 60세; 종양 크기:65개 사례 ≥ 5 cm 및 45개 사례 <5cm; 림프절 전이(LNM)가 없는 경우 71건 및 LNM이 있는 경우 39건; 종양, 결절 및 전이(TNM) 병기:I + II 병기에 60건, III 병기에 50건이 있었습니다. 종양 위치:13건은 상부 ESCC였고 97건은 중하부 ESCC였다. 환자들은 모두 ESCC 진단을 받았고 이전에 방사선 요법이나 화학 요법을 받은 적이 없었습니다. 종양은 완전히 절제되었고, 병리학에서 수술적 절제면이 음성으로 확인되었습니다. 2009년 UICC(Union for International Cancer Control)에서 제안한 ESCC의 병기 기준[14]에 따르면 환자의 수술 후 병리학적 병기는 pT1-4N1-2(I-IIIb)기로 확인되었다. 수술 후 환자에서 유의한 합병증은 없었으며 수술 전후 사망은 제외되었습니다.
역전사 정량 중합효소 연쇄 반응(RT-qPCR)
조직 및 세포의 총 RNA는 Trizol 키트(Invitrogen Inc., Carlsbad, CA, USA)를 사용하여 추출되었습니다. RNA 농도와 품질을 측정했습니다. RNA 프라이머(표 1)는 TaKaRa Biotechnology Co., Ltd.(Liaoning China)에 의해 설계 및 합성되었습니다. RNA는 g DNA Eraser(Takara)가 포함된 Takara PrimeScript™ RT 시약 키트의 지침에 따라 cDNA로 역전사되었습니다. Power PCR SYBR 녹색 PCR 마스터 믹스(Takara)를 사용하여 Light Cycler 480II(Roche)에서 qPCR을 수행했습니다. U6은 miR-301의 로딩 컨트롤로 사용되었으며 β-액틴은 PTEN의 내부 참조로 사용되었습니다. 2
−△△Ct
를 이용하여 데이터 분석 방법 [15].
서부 얼룩 분석
RIPA 용해 완충액(Beyotime Institute of Biotechnology, Shanghai, China)을 사용하여 세포 및 조직의 총 단백질을 추출하고 BCA Protein Assay 키트(Beyotime)로 단백질을 정량하였다. 각 시료의 단백질 농도를 측정하고 10% sodium dodecyl sulfate-polyacrylamide gel 전기영동을 하였다. 샘플을 니트로셀룰로오스 멤브레인으로 옮기고 4°C에서 밤새 5% 탈지분유로 차단했습니다. 그 후, 막을 밤새 배양하기 위해 1차 항체 PTEN 및 β-액틴(둘 다 1:500 및 Santa Cruz Biotechnology Inc, CA, USA)으로 보충한 다음, 각각의 2차 항체를 첨가하고 1시간 동안 배양하였다. 강화된 화학발광 시약(Pierce Chemical Inc., Dallas, TX, USA)에 1분 동안 담근 후, 멤브레인을 어두운 환경에 노출시키고 LAS4000 미니 화학발광 이미저를 사용하여 현상했습니다. 회색 값은 대조군으로 β-액틴을 사용하는 이미징 시스템 소프트웨어에 의해 평가되었습니다. 따라서 발현된 최종 상대 단백질이 얻어졌다. ImageJ2x 소프트웨어로 단백질 밴드를 분석했습니다.
이중 루시퍼라제 리포터 유전자 분석
PTEN의 3'-untranslated region(UTR) 서열은 miR-301과 상호작용할 것으로 예측되거나, 예측된 표적 부위 내의 돌연변이된 서열이 합성되어 pGL3 대조군 루시페라제 리포터 벡터(Promega, 위스콘신, 미국). 그 다음, pGL3-PTEN-wt 및 pGL3-PTEN-mut 벡터를 생산하였다. miR-301 모방체 및 모방체 NC가 있는 올바르게 식별된 wt 및 mut 루시퍼라제 리포터 플라스미드를 48시간 동안 KYSE30 및 Eca109 세포에 공동 형질감염시켰다. 이어서, 세포를 용해시키고, 루시퍼라제 검출 키트(Promega)를 이용하여 각각 루시퍼라제 활성을 측정하였다.
세포 배양, 그룹화 및 형질감염
ESCC 세포주(KYSE-150, KYSE-30, Eca109 및 KYSE-70)는 Shanghai Institute of Biochemistry and Cell Biology, Chinese Academy of Sciences(Shanghai, China)에서 획득했으며 인간 식도 상피 세포(HEEC)는 획득했습니다. Mingzhou Biotechnology Co., Ltd. (Zhejiang, China). 세포를 10% 소 태아 혈청(FBS, Life Technologies, USA), 100단위/ml 페니실린 G 나트륨(Sigma) 및 100㎍/ml 스트렙토마이신 설페이트(Sigma)가 보충된 RPMI 1640 배지(Invitrogen)에서 배양했습니다. 각 세포주에서 MiR-301 발현과 PTEN의 mRNA 발현을 RT-qPCR로 측정하였고, 이후의 세포 실험을 위해 상대 발현이 가장 높고 가장 낮은 세포주를 선택하였다.
KYSE-30 세포를 7개 그룹으로 분리하고 각각 miR-301 억제제, 억제제 음성 대조군(NC), pcDNA-PTEN(과발현(oe)-PTEN으로 명명), pcDNA-NC(oe-NC로 명명), miR-301 억제제 + 작은 간섭 RNA(si)-PTEN 또는 miR-301 억제제 + si-NC. Eca109 세포도 7개의 그룹으로 분리되었고 miR-301 모방체, 모방체 NC, si-PTEN, si-NC, miR-301 모방체 + oe-PTEN, miR-301 모방체 + oe-NC로 여러 번 처리되었습니다. 억제제 NC, miR-301 억제제, miR-301 모방체, 모방체 NC, si-NC 및 si-PTEN은 GenePharma Ltd., Company(중국 상하이)에서 구입했습니다. pcDNA-PTEN NC 및 pcDNA-PTEN은 (Shanghai Sangon Bio-technology Corporation (Shanghai, China))에서 입수했습니다. 세포 융합이 60%에 도달했을 때 세포를 lipofectamine 2000(Invitrogen)에 의해 ESCC 세포에 일시적으로 형질감염시켰습니다.
세포 계수 키트(CCK-8) 분석
세포를 96웰 플레이트(1 × 10
3
세포/웰) 및 다양한 기간 동안 인큐베이션. 24시간, 48시간, 72시간, 96시간 배양 후 각 웰에 10 μL CCK-8 용액(5 mg/mL)을 보충한 후, 각 군의 세포를 빛 없이 37 °C에서 배양하였다. 2시간 노출. 450 nm에서의 광학 밀도(OD) 값은 마이크로플레이트 리더(Bio-Rad Laboratories, Hercules, CA, USA)로 분석되었습니다.
집락 형성 분석
형질감염 후 세포를 6웰 플레이트에 500개 세포/웰로 시딩하고 14일 동안 배양하였다. 콜로니를 메탄올로 고정하고 0.5% 크리스탈 바이올렛을 사용하여 염색하고 도립 현미경으로 계수했습니다.
트랜스웰 분석
셀(5 × 10
3
) ) RPMI 1640 배지에서 배양된 배지를 코팅되지 않은 또는 마트리겔 코팅된 멤브레인(Corning, NY, USA)이 있는 Transwell 장치의 정점 챔버에 접종했습니다. 24시간 후, apical chamber의 세포를 제거하고 아래쪽에 남아 있는 세포를 고정한 다음 0.1% crystal violet을 사용하여 염색하였다. 현미경(Olympus Corporation, Tokyo, Japan)을 사용하여 3개의 무작위 필드를 계산하여 세포 수를 계산했습니다.
유세포분석
세포 주기 및 세포자멸사는 유세포 분석에 의해 평가되었습니다. Annexin V-fluorescein isothiocyanate(10 µL) 및 propidium iodide(PI; 5 µL, Sigma)를 세포와 함께 배양했습니다(5 × 10
5
세포/웰) 30분 동안 4°C에서 어둠 속에서. 세포 사멸 세포의 백분율은 FlowJo 버전 10 소프트웨어(FlowJo LLC, OR, USA)와 함께 유세포 분석기(BD Biosciences, CA, USA)를 사용하여 계산되었습니다.
세포 주기를 평가하기 위해 세포(5 × 10
5
세포/웰)을 4°C에서 밤새 75% 에탄올로 고정하고 어둠 속에서 30분 동안 4°C에서 5 μl PI/리보뉴클레아제 A(Sigma)로 염색했습니다. 데이터는 유세포 분석기(BD Biosciences)로 분석되었습니다. ModFit LT 버전 3.2 소프트웨어(Verity Software House, Inc., ME, USA)를 사용하여 각 샘플의 형광 신호(14,000)를 수집하고 계산했습니다.
누드 마우스의 피하 종양 발생
42마리의 암컷 BALB/c-nu 누드 마우스(4w 노화, 16-24g 무게)를 실험 동물 센터 Jilin University(중국 창춘)에서 얻었다. 누드 마우스는 14개 그룹(n =3). 7개의 그룹의 누드 마우스에는 세포 그룹에 따라 각각 KYSE-30 세포를 주입하고, 나머지 7개 그룹의 누드 마우스에는 그룹에 따라 Eca109 세포를 개별적으로 주입하였다. 형질감염된 KYSE-30 및 Eca109 세포의 농도를 5 × 10
6
으로 조정했습니다. 세포/100 μL. 누드 마우스를 고정하고 멸균 조건에서 상응하는 ESCC 세포를 피하 주사하였다. 매주 종양의 가장 큰 길이(L)와 너비(W)를 측정하고 종양 부피(V ) =1/2 × L × 와
2
. 누드 마우스는 5
에 안락사되었습니다. 종양을 주사한 주에 절제하고 종양의 무게를 측정하고 사진을 찍었습니다. 종양 형성 비율은 피하 종양이 있는 마우스의 수/그룹에서 주사된 누드 마우스의 총 수 × 100%로 계산되었습니다. 주사 시간을 횡축으로, 종양 크기를 종축으로 하였다. 따라서 종양 성장 곡선을 그래프로 표시했습니다.
면역조직화학 염색
누드 마우스의 종양 조직은 10% 포름알데히드로 고정되었고, 파라핀으로 포매되어 4μm로 절단되었습니다. 다음으로, 섹션을 60°C에서 2시간 동안 건조하고, 크실렌으로 탈랍하고, 구배 에탄올로 탈수하고, 50μL 3% H2와 함께 인큐베이션했습니다. O2 10분 동안 그 후, 절편을 0.01M 시트르산 완충액에 담그고 95°C에서 20분 동안 끓인 다음 37°C에서 10분 동안 일반 염소 혈청 작업 용액으로 차단하고 CD34를 추가했습니다. (1:100, Santa Cruz) 4°C에서 밤새. 그 후, 섹션을 HRP로 표지된 염소 항토끼/마우스 IgG 폴리머(ZSGB-Bio, Beijing, China)로 보충하고, 헤마톡실린으로 대조염색하고, 탈수 및 투과화한 다음, 중성 발삼으로 밀봉했습니다. PBS는 NC로 1차 항체를 대체하는 데 사용되었습니다. 미세혈관 밀도(MVD) 측정:절편을 저배율 현미경으로 관찰했습니다. 내피세포 또는 내피세포 군집은 황갈색으로 염색되어 주변 종양세포와 현저하게 구별되며, 결합조직은 미세혈관으로 하였다. 분지 구조는 연결이 끊어진 경우 혈관으로 간주되었으며 내강 크기가> 8 적혈구, 근육층 또는 더 두꺼운 내강을 가진 혈관은 제외되었습니다. 3개의 높은 시야의 미세혈관의 수를 기록하였고, 각 경우의 평균 수는 MVD였다.
통계 분석
모든 통계 분석은 SPSS 버전 SPSS 21.0 소프트웨어(IBM Corp. Armonk, NY, USA)를 사용하여 수행되었으며 Graphpad Prism Software 6.0과 함께 제공되었습니다. 데이터는 평균 ± 표준편차로 표현하였다. 두 개의 독립 그룹 간의 차이는 스튜던트 t 테스트로 테스트되었습니다. 3개 이상의 그룹을 비교하기 위해 일원 ANOVA를 수행했습니다. 피 value <0.05는 통계적으로 유의한 차이를 나타냅니다.
<섹션 데이터-제목="결과">
결과
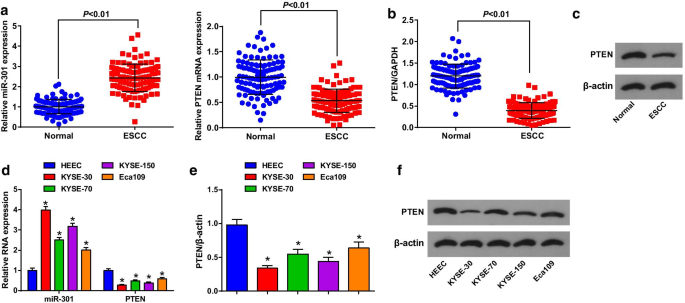
MiR-301은 고도로 발현되지만 PTEN은 ESCC 조직 및 세포에서 제대로 발현되지 않습니다.
ESCC 조직 및 인접 정상 조직에서의 MiR-301 및 PTEN 발현은 RT-qPCR 및 Western blot 분석을 사용하여 ESCC에서의 역할을 밝히기 위해 평가되었으며 (그림 1a-c) miR-301이 상향 조절되는 반면 PTEN ESCC 조직에서 하향조절되었다. miR-301 또는 PTEN 발현의 중앙값에 따라 환자를 저발현군과 고발현군으로 나누어 ESCC 환자의 miR-301 또는 PTEN 발현과 임상병리학적 특성 간의 상관관계를 분석하였다. 결과는 miR-301/PTEN 발현이 연령, 성별, 종양 크기, 위치 및 분화와는 관련이 없는 반면, ESCC 환자의 TNM 병기 및 LNM과 상관관계가 있음을 나타냈다(표 2).
<그림>
MiR-301은 고도로 발현되는 반면 PTEN은 ESCC 조직 및 세포에서 제대로 발현되지 않습니다. 아 RT-qPCR을 사용하여 검출된 ESCC 조직에서 miR-301의 발현 및 PTEN의 mRNA 발현; ㄴ 웨스턴 블롯 분석을 사용하여 검출된 ESCC 조직에서 PTEN의 단백질 발현; ㄷ 웨스턴 블롯 분석에서 ESCC 조직에서 PTEN의 단백질 밴드; d RT-qPCR을 사용하여 검출된 ESCC 세포주에서 miR-301의 발현 및 PTEN의 mRNA 발현; 이 웨스턴 블롯 분석을 사용하여 검출된 ESCC 세포주에서 PTEN의 단백질 발현; 에 웨스턴 블롯 분석에서 PTEN의 단백질 밴드. *피 <0.05 대 HEEC. 데이터는 평균 ± 표준편차로 표현하였으며, 두 집단간 비교를 위해 t검정을 시행하였다
그런 다음, 4개의 ESCC 세포주 및 HEEC에서 miR-301 및 PTEN의 발현을 RT-qPCR 및 웨스턴 블롯 분석을 사용하여 결정하였다. 우리는 (그림 1d-f) miR-301이 ESCC 세포주에서 상향 조절되고 PTEN이 하향 조절된다는 것을 발견했습니다. 그 중 KYSE-30은 가장 높은 miR-301 발현과 가장 낮은 PTEN 발현을 가졌지만 Eca109는 반대 경향을 보였습니다. 따라서 세포 실험에서 KYSE-30 세포주는 하향조절된 miR-301/과발현된 PTEN으로 처리되었고 Eca109 세포주는 과발현된 miR-301/침묵된 PTEN으로 처리되었다.
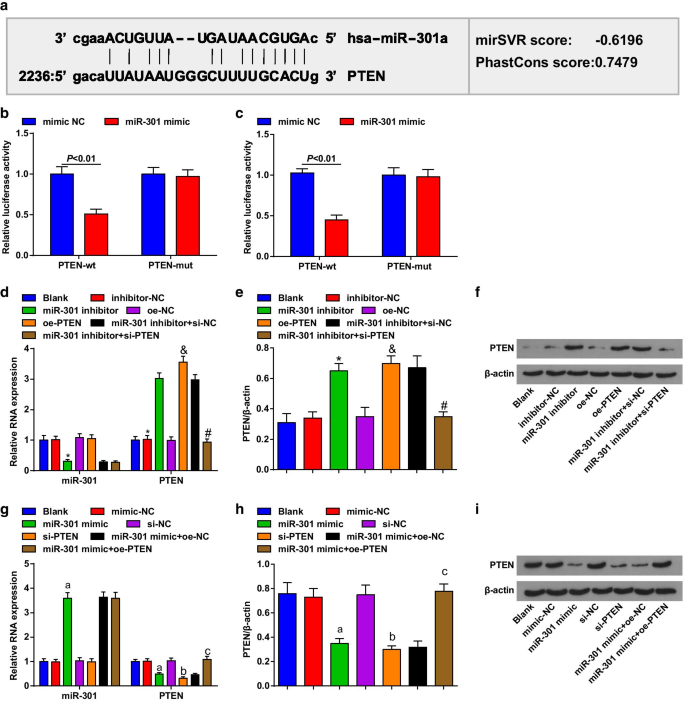
PTEN은 miR-301의 표적입니다
생물정보 소프트웨어(http://www.microrna.org/)는 PTEN이 miR-301의 표적 유전자라고 예측했습니다(그림 2a). 이중 루시페라제 리포터 유전자 분석에 의해 PTEN-mut 벡터 및 miR-301 모방체로 동시 형질감염된 세포와 비교하여 PTEN-wt 벡터 및 miR-301 모방체로 동시 형질감염된 ESCC 세포에서 루시페라아제 활성이 유의하게 감소되었음을 추가로 확인하였다. miR-301이 특히 PTEN에 결합할 수 있음을 암시합니다(그림 2b, c).
<그림>
PTEN은 miR-301의 표적 유전자입니다. 아 miR-301 및 PTEN의 결합 부위는 온라인 예측 소프트웨어에 의해 예측되었습니다. ㄴ KYSE-30 세포에서 miR-301과 PTEN 사이의 표적 관계는 이중 루시퍼라제 리포터 유전자 분석에 의해 평가되었습니다. ㄷ Eca109 세포에서 miR-301과 PTEN 사이의 표적 관계는 이중 루시퍼라제 리포터 유전자 분석에 의해 평가되었습니다. d miR-310 하향 조절 또는 PTEN 상향 조절 후 RT-qPCR을 사용하여 검출된 KYSE-30 세포에서 miR-301 및 PTEN mRNA 발현의 발현; 이 miR-310 하향 조절 또는 PTEN 상향 조절 후 웨스턴 블롯 분석을 사용하여 검출된 KYSE-30 세포에서 PTEN의 단백질 발현; 에 miR-310 하향 조절 또는 PTEN 상향 조절 후 웨스턴 블롯 분석에서 KYSE-30 세포에서 PTEN의 단백질 밴드; 지 miR-310 상향 조절 또는 PTEN 하향 조절 후 RT-qPCR을 사용하여 검출된 Eca109 세포에서 miR-301의 발현 및 PTEN의 mRNA 발현; 아 miR-310 상향 조절 또는 PTEN 하향 조절 후 웨스턴 블롯 분석을 사용하여 검출된 Eca109 세포에서 PTEN의 단백질 발현; 나 miR-310 상향 조절 또는 PTEN 하향 조절 후 웨스턴 블롯 분석에서 Eca109 세포의 PTEN 단백질 밴드. *피 <0.05 대 억제제-NC 그룹, &P <0.05 대 oe-NC 그룹,
#피 <0.05 대 miR-301 억제제 + si-NC 그룹, a P <0.05 대 모방-NC 그룹, b P <0.05 대 si-NC 그룹, c P <0.05 대 miR-301 모방 + oe-NC 그룹; 아니 =3. 자료는 평균 ± 표준편차로 표현하였으며, 두 집단간의 비교를 위해 t검정을 시행하였다. ANOVA는 여러 그룹 간의 비교에 사용되었습니다.
RT-qPCR 및 웨스턴 블롯 분석을 사용하여 형질 감염된 세포에서 miR-301 및 PTEN 발현을 평가했으며 KYSE-30 세포 (그림 2d-f)에서 miR-301 억제제로 처리 된 세포가 miR-301을 하향 조절하는 것으로 나타났습니다 , 상향조절된 PTEN; pcDNA-PTEN(oe-PTEN)으로 처리된 세포는 PTEN 발현을 증가시켰고, si-PTEN은 miR-301 억제제가 PTEN 발현에 미치는 영향을 역전시켰습니다. Eca109 세포(그림 2g-i)에서 miR-301로 처리된 세포는 miR-301을 상향 조절한 반면 PTEN을 하향 조절했습니다. si-PTEN으로 처리된 세포는 PTEN 발현을 감소시켰고 pcDNA-PTEN(oe-PTEN)은 PTEN 발현에서 miR-301 모방의 억제 역할을 역전시켰습니다. 이 데이터는 miR-301이 PTEN을 표적으로 했음을 시사합니다.
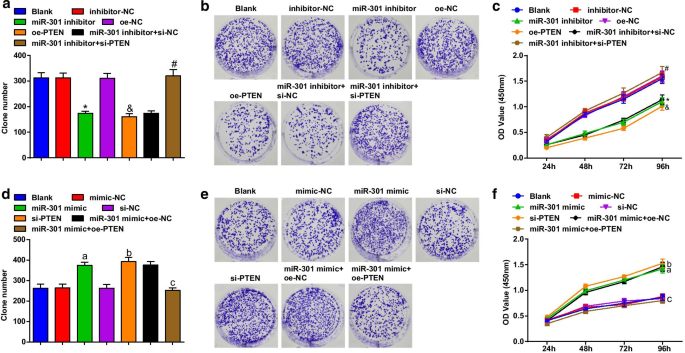
억제된 miR-301 또는 과발현된 PTEN은 ESCC 세포의 생존을 제한합니다. 증가된 miR-301 또는 감소된 PTEN은 ESCC 세포의 생존을 촉진합니다.
ESCC 세포의 세포 생존력은 콜로니 형성 및 CCK-8 분석을 사용하여 평가되었습니다. 결과는 KYSE-30 세포주(그림 3a-c)에서 miR-301 억제제 또는 oe-PTEN의 형질감염이 집락 형성 능력과 세포 생존 능력을 억제한다는 것을 보여주었습니다. 침묵 PTEN의 형질 감염은 ESCC 세포 생존력에 대한 miR-301 억제제의 영향을 제거했습니다. Eca109 세포주(그림 3d-f)에서 miR-301 모방체 또는 si-PTEN의 형질감염은 집락 형성 능력과 세포 생존력을 촉진했습니다. PTEN 과발현은 Eca109 세포의 집락 형성 능력과 생존력에서 miR-301 상승의 촉진 역할을 역전시켰습니다. 이러한 결과는 miR-301 녹다운 또는 PTEN 과발현이 miR-301 상승 또는 PTEN 억제에 의해 촉진되는 ESCC 세포의 생존력을 억제함을 시사합니다.
<그림>
억제된 miR-301 또는 과발현된 PTEN은 ESCC 세포의 생존을 제한합니다. 증가된 miR-301 또는 감소된 PTEN은 ESCC 세포의 생존을 촉진합니다. 아 miR-310 하향 조절 또는 PTEN 상향 조절 후 콜로니 형성 분석을 사용하여 검출된 형질감염 후 KYSE-30 세포의 콜로니 수; ㄴ miR-310 하향 조절 또는 PTEN 상향 조절 후 집락 형성 분석을 사용하여 검출된 형질감염 후 KYSE-30 세포의 집락 형성 능력; ㄷ miR-310 하향 조절 또는 PTEN 상향 조절 후 CCK-8 분석을 사용하여 검출된 형질감염 후 KYSE-30 세포의 생존력; d miR-310 상향 조절 또는 PTEN 하향 조절 후 콜로니 형성 분석을 사용하여 검출된 형질감염 후 Eca109 세포의 콜로니 수; 이 miR-310 상향 조절 또는 PTEN 하향 조절 후 집락 형성 분석을 사용하여 검출된 형질감염 후 Eca109 세포의 집락 형성 능력; 에 miR-310 상향 조절 또는 PTEN 하향 조절 후 CCK-8 분석을 사용하여 검출된 형질감염 후 Eca109 세포의 생존력; *피 억제제-NC 그룹에 비해 <0.05; &피 <0.05 대 oe-NC 그룹;
#피 <0.05 대 miR-301 억제제 + si-NC 그룹; 피 <0.05 대 모방-NC 그룹; b 피 <0.05 대 si-NC 그룹; c 피 <0.05 대 miR-301 모방 + oe-NC 그룹, N =3. 데이터는 평균 ± 표준편차로 표현하였으며, 여러 그룹간 비교는 ANOVA를 이용하였다
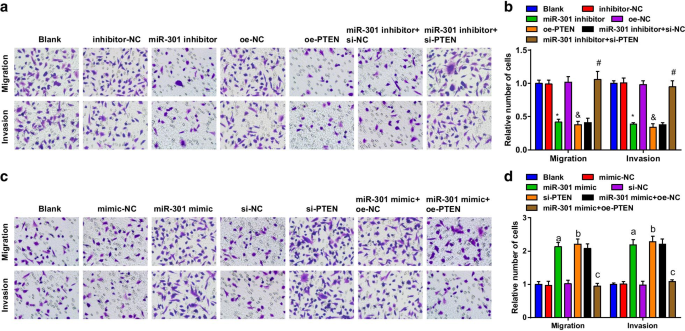
억제된 miR-301 또는 과발현된 PTEN은 ESCC 세포의 이동 및 침입을 억제합니다. 증가된 miR-301 또는 감소된 PTEN은 ESCC 세포의 이동 및 침범을 유도합니다.
Transwell 분석을 사용하여 ESCC 세포의 이동 및 침입 능력을 평가했습니다. 결과는 KYSE-30 세포주(그림 4a, b)에서 miR-301의 억제 또는 PTEN의 과발현에 의해 세포 이동 및 침입 능력이 억제되었음을 시사하였다. 세포 이동 및 침입 능력에서 miR-301 억제제의 억제 역할은 si-PTEN에 의해 역전되었습니다. Eca109 세포주(도 4c, d)에서 miR-301 모방체 또는 si-PTEN의 형질감염 후 세포 이동 및 침입 능력이 촉진되었다. 과발현된 PTEN은 세포 이동 및 침입 능력에 대한 miR-301 모방의 영향을 역전시켰습니다. 위의 결과는 ESCC 세포의 이동 및 침범이 miR-301 억제 또는 PTEN 상승에 의해 억제되는 반면 miR-301 상향 조절 또는 PTEN 하향 조절에 의해 촉진됨을 의미합니다.
<그림>
억제된 miR-301 또는 과발현된 PTEN은 ESCC 세포의 이동 및 침입을 억제합니다. 증가된 miR-301 또는 감소된 PTEN은 ESCC 세포의 이동 및 침입을 촉진합니다. 아 miR-310 하향 조절 또는 PTEN 상향 조절 후 Transwell 분석을 사용하여 평가된 형질감염된 KYSE-30 세포의 이동 및 침입 능력; ㄴ miR-310 하향 조절 또는 PTEN 상향 조절 후 Transwell 분석을 통한 KYSE-30 세포의 이동 및 침습의 통계적 결과; ㄷ miR-310 상향 조절 또는 PTEN 하향 조절 후 Transwell 분석을 사용하여 평가된 그룹 중 Eca109 세포의 이동 및 침입 능력; d Transwell assay miR-310 upregulation 또는 PTEN downregulation을 통한 Eca109 세포의 이동 및 침습에 대한 통계적 결과. *피 억제제-NC 그룹에 비해 <0.05; &피 <0.05 대 oe-NC 그룹;
#피 <0.05 대 miR-301 억제제 + si-NC 그룹; 피 <0.05 대 모방-NC 그룹; b 피 <0.05 대 si-NC 그룹; c 피 <0.05 대 miR-301 모방 + oe-NC 그룹, N =3. 데이터는 평균 ± 표준편차로 표현하였으며, 여러 그룹간 비교는 ANOVA를 이용하였다
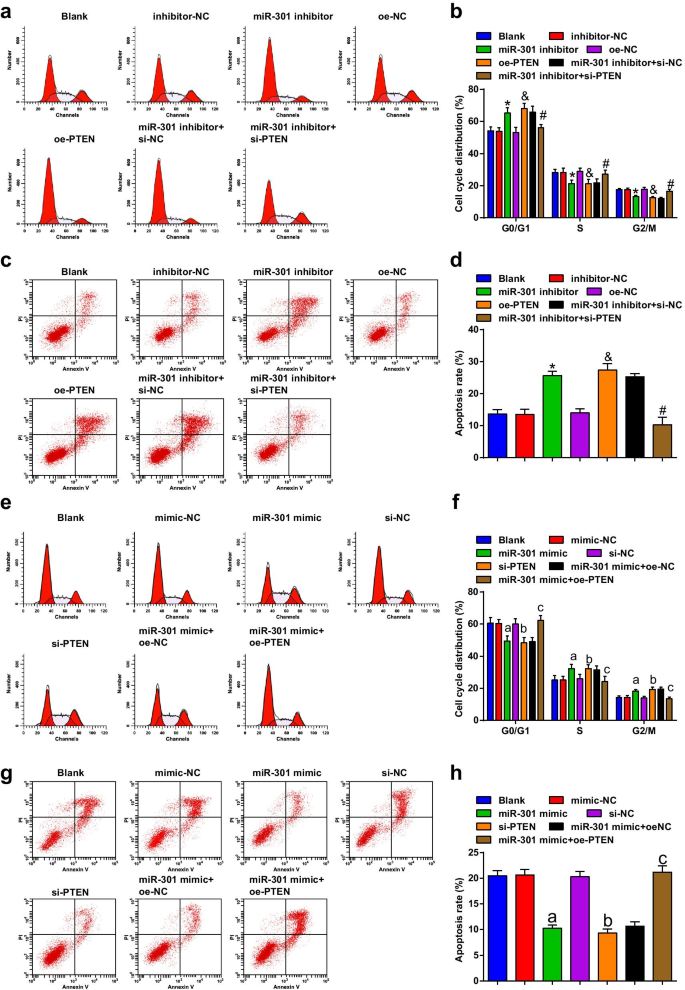
억제된 miR-301 또는 과발현된 PTEN은 ESCC 세포의 세포 주기 정지 및 아폽토시스를 유도합니다. 증가된 miR-301 또는 감소된 PTEN은 ESCC 세포의 세포 주기 정지 및 아폽토시스를 억제합니다.
Flow cytometry는 transfection 후 cell의 cell cycle transition과 apoptosis를 감지하는데 사용되었으며, 결과는 KYSE-30 cell line(Fig. 5a-d)에서 miR-301 inhibitor나 oe-PTEN의 transfection이 apoptotic rate를 증가시키는 것으로 나타났다. G0/G1기에서는 세포가 증가했지만 S 및 G2/M기에서는 세포가 감소했습니다. miR-301 억제제에 의해 유도된 세포 사멸 및 세포 주기 정지의 변경은 si-PTEN에 의해 역전될 수 있습니다.
<그림>
억제된 miR-301 또는 과발현된 PTEN은 ESCC 세포의 세포 주기 정지 및 세포자멸사를 유도합니다. 증가된 miR-301 또는 감소된 PTEN은 ESCC 세포의 세포 주기 정지 및 세포 사멸을 억제합니다. 아 각 그룹에서 KYSE-30 세포의 세포 주기 분포는 miR-310 하향 조절 또는 PTEN 상향 조절 후 유세포 분석에 의해 검출되었습니다. ㄴ miR-310 하향 조절 또는 PTEN 상향 조절 후 유세포 분석에서 KYSE-30 세포의 G0/G1, S 및 G2/GM 단계의 백분율 통계 결과; ㄷ KYSE-30 세포의 아폽토시스는 miR-310 하향 조절 또는 PTEN 상향 조절 후 유세포 분석에 의해 감지되었습니다. d miR-310 하향 조절 또는 PTEN 상향 조절 후 유세포 분석을 사용하여 검출된 형질감염된 KYSE-30 세포의 세포자멸사 비율; 이 각 그룹에서 Eca109 세포의 세포 주기 분포는 유세포 분석 miR-310 상향 조절 또는 PTEN 하향 조절에 의해 검출되었습니다. 에 miR-310 상향 조절 또는 PTEN 하향 조절 후 유세포 분석에서 Eca109 세포의 G0/G1, S 및 G2/GM 단계의 백분율 통계 결과; 지 Eca109 세포의 세포 사멸은 miR-310 상향 조절 또는 PTEN 하향 조절 후 유세포 분석에 의해 감지되었습니다. 아 miR-310 상향 조절 또는 PTEN 하향 조절 후 유세포 분석을 사용하여 검출된 형질감염된 Eca109 세포의 세포자멸사 속도. *피 억제제-NC 그룹에 비해 <0.05; &피 <0.05 대 oe-NC 그룹;
#피 <0.05 대 miR-301 억제제 + si-NC 그룹; 피 <0.05 대 모방-NC 그룹; b 피 <0.05 대 si-NC 그룹; c 피 <0.05 대 miR-301 모방 + oe-NC 그룹, N =3. 데이터는 평균 ± 표준편차로 표현하였으며, 여러 그룹간 비교는 ANOVA를 이용하였다
유세포 분석의 결과에 따르면, 우리는 Eca109 세포주(그림 5e-h)에서 miR-301 모방체 또는 si-PTEN의 형질 감염이 세포 사멸 속도를 억제하고 G0/G1 단계에서 세포를 감소시키고 세포 S상 및 G2/M상; PTEN 과발현은 Eca109 세포의 세포사멸 속도와 세포 주기 정지에 대한 miR-301 모방의 효과를 역전시켰습니다. 우리는 이러한 결과로부터 miR-301을 하향 조절하거나 PTEN을 상향 조절하면 ESCC 세포에서 세포 주기 전환 및 세포자멸사를 촉진하는 반면 miR-301을 억제하거나 PTEN을 침묵시키는 것은 반대 효과를 발휘한다는 결론을 내렸습니다.
억제된 miR-301 또는 과발현된 PTEN은 ESCC에서 생체내 종양 성장 및 혈관신생을 억제합니다. 증가된 miR-301 또는 감소된 PTEN은 ESCC에서 생체 내 종양 성장 및 혈관신생을 증가시킵니다.
누드 마우스에서 ESCC 종양의 성장과 변화는 각 그룹에서 관찰되었습니다. 종양 성장을 평가했으며 결과는 KYSE-30 세포주(그림 6a-e)에서 miR-301 억제제 또는 oe-PTEN을 주사한 누드 마우스가 종양 부피와 무게를 감소시켰음을 암시했습니다. 종양 성장에서 miR-301 억제제의 억제 역할은 si-PTEN에 의해 폐지되었습니다. Eca109 세포주(그림 6f-j)에서 miR-301 모방체 또는 si-PTEN을 주사한 누드 마우스에서 종양 부피와 종양 무게가 진행되었습니다. PTEN의 과발현은 종양 성장에 대한 miR-301 모방의 효과를 역전시켰습니다. 한편, 누드 마우스의 이종이식편에서 CD34의 발현은 면역조직화학적 염색을 사용하여 평가되었고 그 결과는 KYSE-30 이종이식편에서 (그림 7a-d) miR-301 하향 조절 또는 PTEN 상향 조절 후에 MVD가 억제됨을 보여주었습니다. 침묵 PTEN은 MVD에 대한 miR-301 억제의 영향을 역전시켰습니다. Eca109 이종이식편에서 miR-301 상향 조절 또는 PTEN 하향 조절 후 MVD가 증가했습니다. 상향 조절된 miR-301에 의해 유도된 MVD의 향상은 과발현된 PTEN에 의해 폐지될 수 있습니다. 이러한 데이터는 miR-301 억제 또는 PTEN 과발현이 ESCC에서 종양 성장 및 혈관신생을 억제하는 반면 miR-301 상승 또는 PTEN 침묵은 역효과를 갖는다는 것을 나타냅니다.
Inhibited miR-301 or overexpressed PTEN restrains tumor growth in ESCC; elevated miR-301 or reduced PTEN increases tumor growth in ESCC. 아 Representative figures for the tumor growth observed by subcutaneous tumorigenesis in nude mice after KYSE-30 cells were transfected; b–d changes of tumor volume of each group after KYSE-30 cells were transfected; 이 changes of tumor weight of each group after KYSE-30 cells were transfected; 에 representative figures for the tumor growth observed by subcutaneous tumorigenesis in nude mice after Eca109 cells were transfected; g–i changes of tumor volume of each group after Eca109 cells were transfected; j changes of tumor weight of each group after Eca109 cells were transfected. *피 < 0.05 versus the inhibitor-NC group; &피 < 0.05 versus the oe-NC group;
#피 < 0.05 versus the miR-301 inhibitors + si-NC group; a P < 0.05 versus the mimic-NC group; b P < 0.05 versus the si-NC group; c P < 0.05 versus the miR-301 mimic + oe-NC group, n = 3 mice. Data were expressed as mean ± standard deviation, and ANOVA was used for comparisons among multiple groups
Inhibited miR-301 or overexpressed PTEN restrains angiogenesis in ESCC; elevated miR-301 or reduced PTEN increases angiogenesis in ESCC. 아 Representative images of tumor tissues observed by immunohistochemical staining in nude mice after KYSE-30 cells were transfected; ㄴ comparisons of MVD of KYSE-30 in tumor tissues among the groups; ㄷ representative images of tumor tissues observed by immunohistochemical staining in nude mice after Eca109 cells were transfected; d comparisons of MVD of Eca109 in tumor tissues among the groups *P < 0.05 versus the inhibitor-NC group; &피 < 0.05 versus the oe-NC group;
#피 < 0.05 versus the miR-301 inhibitors + si-NC group; a P < 0.05 versus the mimic-NC group; b P < 0.05 versus the si-NC group; c P < 0.05 versus the miR-301 mimic + oe-NC group, n = 3 mice. Data were expressed as mean ± standard deviation, and ANOVA was used for comparisons among multiple groups
토론
EC is a kind of invasive malignancy in the gastrointestinal tract [16]. As the major type of EC, ESCC is a malignant tumor occurring in esophageal epithelial cells [17]. The miRNAs, known as small non-coding RNAs, have been demonstrated to function as a significant roles in leading molecules in the silencing of RNA [18]. Our research was designed to explore the effects of miR-301 and its target gene PTEN on ESCC progression, and we have found that the inhibited miR-301 could suppress angiogenesis and cell growth in ESCC by elevating PTEN.
MiR-301 expression was assessed, and we found that miR-301 was highly expressed in ESCC cells in comparison with HEEC, and the higher expression of miR-301 has also been found in ESCC tissues in contrast to the adjacent normal tissues. Similar to this result, Li et al. have identified that miR-301 presented high expression in myocardial infarction tissues [19]. In addition, we have elucidated that PTEN was targeted by miR-301, and the target relation has been pointed out by an extant literature [20]. We have also discovered that PTEN, which has been affirmed to be targeted by miR-301, was downregulated in both ESCC tissues and cells. Similarly, a previous research has unearthed that PTEN was poorly expressed in ESCC compared with non-tumor esophageal epithelial tissue [21]. Furthermore, Ma et al. have illuminated that PTEN expression was degraded in Eca109 cell line [22], which has also been selected for a series of experiments in this research. These studies provide evidence for the high expression of miR-301 and low expression of PTEN in ESCC.
Another important outcome in this research indicated that the inhibited miR-301 could repress the colony formation ability as well as the cell proliferation of ESCC cells via enhancing the PTEN expression, and elevated miR-301 or reduced PTEN had contrary effects. Similarly, Han et al. have elucidated that the downregulation of miR-301 mediated by luteolin has the ability to restrain the cell proliferation in prostate cancer [6]. A recent literature has revealed that the overexpression of PTEN suppresses the proliferation of pancreatic cancer cells [23], and a same result has been summarized in a study focusing on prostate cancer [24]. Besides, we have also unearthed that the downregulation of miR-301 or the elevation of PTEN could inhibit migration and invasion of ESCC cells, and elevated miR-301 or reduced PTEN exhibited the opposite trends. In accordance with this outcome, Shi et al. have supported that inhibited miR-301 attenuated migration and invasion of breast cancer cells [10], and it has been reported that the migration and invasion of ESCC cells could be repressed by the inhibition of miR-130b and the elevation of PTEN [25]. These publications helped verifying the oncogenic role of miR-301 and tumor-repressive effect of PTEN in diverse human cancers. Another result in our research was that inhibited miR-301 overexpressed PTEN to promote cell apoptosis and induce cell cycle arrest at the G0/G1 phase in ESCC cells, and elevated miR-301 or reduced PTEN had the inverse results. Similarly, it has been uncovered by a recent literature that activated PTEN induces cell cycle arrest and apoptosis in ESCC [26]. Consistently, Tian et al. have found in their study that the elevation of PTEN inhibited the angiogenesis by reducing the expression of vascular endothelial growth factor in hepatocellular carcinoma [27]. Based on the above data, the roles of miR-301 and PTEN in cell apoptosis and angiogenesis in diverse diseases were further confirmed. Consequently, we concluded that the downregulation of miR-301 could restrain the tumor growth in ESCC through the high expression of PTEN, and the similar conclusion has also been unveiled in breast cancer [10] and prostate cancer [28]. On the contrary, miR-301 elevation or PTEN reduction induced the tumor growth in ESCC. It could be concluded that miR-301 and PTEN participated in the in vivo cancer cell growth.
Conclusion
In this study, we have shown that the repression of miR-301 prohibits angiogenesis, cell proliferation, migration and invasion but promotes apoptosis in ESCC cells by upregulating PTEN. This research may further the understanding on potential molecular mechanisms of ESCC and provide novel targets for ESCC treatment.