SOX2는 lncRNA CCAT1/MicroRNA-185-3p/FOXP3 축을 조절하여 자궁경부암 줄기 세포의 증식 및 자가 재생에 영향을 줍니다.
초록
자궁경부암(CC)에서 긴 비암호화 RNA(lncRNA)의 역할이 제시되었습니다. 우리는 성 결정 영역 Y-box 2(SOX2)/lncRNA 결장암 관련 전사체-1(CCAT1)/microRNA-185-3p(miR-185-3p)/forkhead box protein 3(FOXP3)의 효과에 대해 논의하는 것을 목표로 합니다. ) CC 줄기 세포의 증식 및 자가 재생 능력에 관한 것입니다. MiR-185-3p, SOX2, CCAT1 및 FOXP3 발현은 CC 조직 및 세포에서 테스트되었습니다. CC 환자에서 SOX2/CCAT1 발현과 임상병리학적 특징 사이의 관계가 확인되었습니다. 기능 손실 및 획득 조사는 CD44
+
에서 수행되었습니다. HeLa 세포는 생물학적 기능과 자가 재생 능력에 대해 논의합니다. 마지막으로 SOX2, CCAT1, FOXP3 및 miR-185-3p 간의 관계를 확인했습니다. miR-185-3p 발현은 감소한 반면, SOX2, CCAT1 및 FOXP3 발현은 CC 조직 및 세포에서 증가하였다. SOX2 및 CCAT1 발현은 종양 크기, 림프절 전이 및 CC의 국제 산부인과 연맹 및 산과 단계와 관련이 있었습니다. SOX2 또는 CCAT1을 하향 조절하고 miR-185-3p를 상향 조절하면 CD44
+
의 세포자멸사 촉진뿐만 아니라 증식, 침입, 이동 및 세포구 수의 억제가 발생합니다. 헬라 세포. SOX2는 miR-185-3p 발현에 영향을 미치는 CCAT1에 결합할 수 있으며, FOXP3는 miR-185-3p의 표적이 됩니다.
소개
자궁경부암(CC)은 2018년 전 세계적으로 약 570,000건이 발생하고 311,000명이 사망하는 여성의 네 번째 주요 사망 원인입니다[1]. 이 복잡한 질병은 유전적 영향과 바이러스 감염을 포함한 다양한 요인에 관여합니다[2]. 인유두종바이러스 공동검사와 인유두종바이러스 백신 접종의 발달로 자궁경부 이형성증과 암의 조기 진단 절차는 CC의 발병률, 이환율 및 사망률을 감소시킵니다[3]. 초기 CC 환자의 경우 생식력 보존 수술, 원뿔 생검, 근치 기관 절제술, 골반 림프절 절제술, 골반 방사선 요법 및 근접 치료와 같은 수술이 권장됩니다[4]. 진행성 CC 환자의 전이 또는 재발로 인해 여전히 예후가 좋지 않습니다[5]. 따라서 CC의 치료를 개선하기 위한 새롭고 효과적인 예후 마커 및 치료 전략을 식별하는 것이 여전히 시급합니다.
SOX2(Sex-determining region Y-box 2)는 전사인자 SOX 계열의 중요한 구성원이며 주로 배아 및 성체줄기세포에서 발현되며 종양줄기세포에서도 발현된다[6]. SOX2가 고슴도치 신호 경로에 의해 CC의 방사선 저항을 조절한다는 것이 밝혀졌습니다[7]. 또 다른 연구는 SOX2가 CC 세포주에서 암 줄기 세포의 하위 집단을 유지하는 데 중요하다는 것을 보여주었습니다[8]. 긴 비암호화 RNA(lncRNA)는 길이가 200개 뉴클레오티드인 RNA 분자 클래스입니다[9]. LncRNA 대장암 관련 전사체-1(CCAT1)은 인간 염색체 8q24.21에 위치하며 대장암에서 유전적 돌연변이를 일으키는 "핫스팟"으로 간주됩니다[10]. 연구에 따르면 CCAT1은 CC의 세포 증식과 침입을 촉진한다고 보고되었습니다[11]. Jia et al.에 따르면, CCAT1은 CC 세포의 증식, 이동 및 침입을 극적으로 향상시킵니다[12]. 또한, 또 다른 연구에서는 CCAT1이 microRNA-185-3p(miR-185-3p)를 감소시켜 장 장벽을 파괴함으로써 염증성 장 질환의 악성 정도를 향상시키는 것으로 밝혀졌습니다[13]. MiRNA는 mRNA를 감소시키고 번역을 억제함으로써 유전자의 발현을 역으로 제어할 수 있다[14]. 이전 연구의 시험관 내 실험에서 miR-185-3p가 비인두암 방사선 저항성을 조절한다는 것이 밝혀졌습니다[15]. 또 다른 연구에서는 miR-185가 생체 내 및 시험관 내 난소암 시스플라틴 내성에 관여한다는 것을 암시했습니다[16]. Forkhead box protein 3(FOXP3)는 FOX 단백질 계열에 속하는 전사 인자로, 조절 T(Treg) 세포에서 처음 발견되며 Treg 세포의 유지 및 과정에 중요한 역할을 합니다[17]. 연구에 따르면 FOXP3는 CC의 림프관 신생과 관련이 있습니다[18]. 또 다른 연구에서는 FOXP3의 수준이 국제 부인과 및 산부인과 병기(FIGO 병기) 및 CC의 종양 크기와 극적으로 연관되어 있음을 보여줍니다[19]. 따라서 본 연구에서는 SOX2/CCAT1/miR-185-3p/FOXP3 축이 CC 줄기 세포의 증식 및 자가 재생 능력에 미치는 영향을 조사했습니다.
자료 및 방법
윤리 승인 및 참여 동의
인간과 관련된 실험은 헬싱키 선언에 명시된 원칙에 따라 시행되었습니다. 이 연구는 길림대학교 제1병원 기관심사위원회의 승인을 받았습니다. 모든 참가자는 사전 동의 문서에 서명했습니다.
연구 주제
2016년 12월부터 2018년 12월까지 CC 환자로부터 39개의 CC 조직 및 해당 인접 정상 조직을 채취하여 액체 질소에 보존했습니다. 포함 기준은 다음과 같다:(1) 환자는 경경부 및 경부 운하의 생검, 자궁 경부 도말 세포학, 자궁 경부 요오드 검사, 질 검경 및 경추 절제술의 병리학에 의해 CC로 확인되었습니다. (2) 환자는 수술 2주 전에 방사선 요법과 화학 요법을 받지 않았습니다. 제외 기준은 (1) 방사선 치료 또는 화학 요법을 받고 있는 환자, (2) 샘플 수집에 동의하지 않는 환자, (3) 면역 체계 장애가 있는 환자였습니다.
세포 선택 및 배양
CC 세포주(SiHa, HeLa, CaSki, HCC94 및 C33A) 및 인간 자궁 경부 상피 불멸화 세포주 H8은 Shanghai Bioleaf Biotech Co., Ltd.(중국 상하이)에서 구입했습니다. CC 세포 SiHa, HeLa 및 HCC94는 10% FBS(fetal bovine serum)를 포함하는 고글루코스 Dulbecco's Modified Eagle Medium(DMEM)에서 배양된 반면, CaSki, C33A 및 H8 세포는 10%를 포함하는 Roswell Park Memorial Institute(RPMI)-1640 배지에서 배양되었습니다. % FBS(37℃ 및 5% CO2 ). 2일마다 세포를 분리하여 계대배양하였다.
CC 줄기 세포 분류(CD44
+
헬라 세포)
CC 줄기 세포는 세포 현탁액 배양에 의해 CC 세포주 HeLa로부터 분리되었다. CC 세포 HeLa는 3-5일마다 반정량적으로 교환된 배지와 함께 21일 동안 초저접착 페트리 접시에서 무혈청 Iscove의 Modified Dulbecco's Medium(IMDM)으로 배양되었습니다. 일부 세포는 구체에 현탁되었고, HeLa-구체 형성 세포(SFC)를 얻었다. 구형 세포의 특성을 확인하고 분석했습니다. 세포를 트립신에 의해 분리하고 1 × 10
6
으로 조정했습니다. 세포/mL. 세포에 CD44 항체를 첨가하고 유세포 분석에 의해 분류하였다. CD44가 양성인 HeLa 세포는 HeLa 종양 줄기 세포였고, CD44가 음성인 HeLa 세포는 비줄기 세포였습니다. CC 줄기 세포를 DMEM/F12에서 배양하고 20ng/mL 기본 섬유아세포 성장 인자(bFGF), 20ng/mL 표피 성장 인자(EGF) 및 B27을 첨가했습니다. 배지에는 1% 페니실린과 스트렙토마이신이 포함되어 있습니다[20].
세포 처리
CD44
+
HeLa 세포는 sh-SOX2, sh-SOX2 음성 대조군(NC), sh-CCAT1, sh-CCAT1 NC, miR-185-3p 모방체, 모방체 NC, sh-CCAT1 및 miR-185-3p 억제제뿐만 아니라 sh-CCAT1 및 억제제 NC. 모든 올리고뉴클레오티드 서열은 GenePharma(Shanghai, China)에서 제공했습니다. 트립신에 의해 분리된 세포는 3 × 10
6
이 포함된 6웰 플레이트에 시딩되었습니다. 세포/웰. 60% 합류에 도달하면 세포를 무혈청 배지로 변경하고 1시간 동안 배양했습니다. Transfection은 Lipofectamine 2000 transfection 시약(Invitrogen, Carlsbad, California, USA)에 의해 촉진되었습니다.
역전사 정량적 중합효소 연쇄 반응(RT-qPCR)
조직 및 세포의 RNA는 Trizol(Invitrogen)로 추출했습니다. RNA(1μg)는 몰로니 쥐 백혈병 바이러스 RTase 키트(Invitrogen)에 의해 cDNA로 역전되었습니다. Real-Time PCR 시스템에 cDNA를 추가했습니다. 프라이머는 Shanghai Sangon Biotechnology Co., Ltd.(중국 상하이)에서 고안했습니다(표 1). U6은 miR-185-3p의 로딩 대조군인 반면, SOX2, CCAT1 및 FOXP3의 글리세르알데히드 포스페이트 탈수소효소(GAPDH)는 로딩 대조군이었다. 결과는 2
-ΔΔCt
로 분석되었습니다. 방법.
서부 얼룩 분석
세포와 조직의 총 단백질을 추출했습니다. 단백질 농도는 비신코닌산 키트(AmyJet Scientific, Wuhan, Wuhan, Hubei, China)에 의해 결정되었습니다. 단백질을 로딩 버퍼와 혼합하고 5분 동안 끓인 다음 ice bath와 원심분리를 수행했습니다. 단백질을 10% 나트륨 도데실 설페이트-폴리아크릴아미드 겔 전기영동으로 처리하고 멤브레인으로 옮겼다. 막을 5% 탈지유로 1시간 동안 차단하고 1차 항체 SOX2(1:1000, Jiangsu Rui sitan Co., Ltd., Jiangsu, China), FOXP3(1:1000, Abcam Inc., Cambridge, MA)로 조사했습니다. , USA), GAPDH(1:1000, Cell Signaling Technology, Beverly, MA, USA) 및 양고추냉이 과산화효소로 표지된 이차 항체로 재탐색하고, 방부제 필름으로 덮고 노출시켰다. Bio-rad Gel Doc EZ 이미저(Bio-rad, California, USA)가 개발을 위해 채택되었습니다. 단백질 이미지는 ImageJ2x 소프트웨어로 분석되었습니다.
세포 계수 키트(CCK)-8 분석
CCK-8 분석은 키트(Beyotime, Shanghai, China)로 구현되었습니다. 셀(1 × 10
4
) )을 96-웰 플레이트에 접종하고 인큐베이션했습니다. 0, 24, 48 및 72시간 동안 배양된 세포에 10μL/웰 CCK-8 용액을 첨가하고 1시간 동안 부화했습니다. 광학 밀도 값은 450nm에서 Multiskan Spectrum 전체 파장 마이크로플레이트 판독기로 결정되었습니다. 평균 값을 계산하기 위해 6개의 웰을 취했습니다. 세포 성장 곡선은 세로 좌표로 시간과 세로 좌표로 상대적인 세포 생존율로 플롯되었습니다. 광학 밀도 값은 세포 증식을 나타냅니다.
유세포분석
셀(1 × 10
6
) )를 1500 r/min에서 원심분리하고, 200 μL 결합 완충액으로 현탁하고, 5 μL 요오드화 프로피듐(PI) 및 5 μL Annexin V-플루오레세인 이소티오시아네이트(FITC)와 차례로 인큐베이션하고 400 μL 완충액을 첨가했습니다. 세포 사멸 속도는 유세포 분석기(BD Biosciences, NJ, USA)로 확인했습니다.
스크래치 테스트
형질감염된 세포를 분리하고 세포 현탁액으로 제조하였다. 세포 현탁액(1 × 10
6
세포/mL)를 6웰 플레이트에 접종하고 80-90% 합류까지 배양했습니다. 6웰 플레이트의 가로 및 세로 스크래치는 눈금자를 따라 10μL 무균 피펫 팁으로 균일하게 만들어졌습니다. 원래 배지를 완전 배지로 교체하고 세포를 지속적으로 배양하였다. 세포의 이동은 0시간과 72시간에 현미경으로 관찰되었으며 동일한 위치에서 촬영되었습니다.
트랜스웰 분석
형질감염된 세포를 트립신 처리하고 1000g에서 원심분리하고 DMEM으로 헹구었습니다. 완전히 수화된 Transwell 챔버를 10% FBS-DMEM(600mL/웰)이 포함된 24웰 플레이트에 넣었습니다. 세포 현탁액(5 × 10
5
세포/mL, 200μL)를 Transwell 상부 챔버(1:8 Matrigel 80μL로 코팅됨)에 추가하고, 500μL 20% FBS-DMEM을 하부 챔버에 추가했습니다. 세포를 24시간 동안 계속 배양하고 500μL 4% 파라포름알데히드(PFA)로 고정하고 0.1% 크리스탈 바이올렛 염색 용액으로 염색했습니다. 그런 다음 면봉으로 상부 챔버 표면과 가장자리의 세포를 닦았습니다. 5개의 필드를 무작위로 선택하고 Nikon Eclipse TE2000-S 현미경(Nikon, Japan)으로 세포를 계수했습니다.
구체 형성 분석
세포를 200개 세포/웰을 갖는 무혈청 현탁액 배지를 함유하는 저흡착 6-웰 플레이트에 접종하였다. 2 w 후, 세포의 구체 형성 속도를 Nikon Eclipse TE2000-S 현미경(Nikon)으로 관찰하였고, 구체 형성 속도는 평균 구체 수/씨딩된 세포 수 × 100%로 계산하였다.
염색질 면역침전(ChIP) 분석
ChIP 분석은 ChIP 키트(Upstate, NY, USA)로 작동되었습니다. SOX2(1:1000, Re-stem Biotech, Jiangsu, China) 및 일반 토끼 IgG(12–370, Millipore, USA) 항체를 사용하여 가교 단백질-DNA 복합체를 면역침전시켰다. CD44
+
HeLa 세포를 1% PFA로 고정하고 배양하여 DNA-단백질 가교를 생성했습니다. 그런 다음 초음파로 DNA를 200–300bp의 염색질 조각으로 절단했습니다. 침전된 염색질 DNA를 회수하여 RT-qPCR로 분석하였다.
RNA 풀다운 분석
비오틴 표지 miR-185-3p 야생형(WT) 및 돌연변이형(MUT) 플라스미드(50nM)를 CD44
+
로 형질감염했습니다. 각각 HeLa 세포. 세포는 48시간 형질감염 후 특정 세포 용해물(Ambion, Austin, Texas, USA)로 부화되었습니다. 세포 용해물(50mL)은 하위 포장되었습니다. 잔류 용해물은 RNase가 없는 효모 tRNA(Sigma)로 미리 코팅된 M-280 스트렙타비딘 비드(Sigma, St. Louis, MO, USA)로 부화되었습니다. 그 다음, 세포를 저온 용해물로 2회, 저염 완충제로 3회, 고염 완충제로 1회 세척하였다. 길항 miR-185-3p 프로브가 NC로 설정되었습니다. Trizol로 total RNA를 추출하고 RT-qPCR로 CCAT1 level을 테스트 하였다.
이중 루시퍼라제 리포터 유전자 분석
CCAT1 프로모터 영역에서 SOX2의 잠재적 결합 부위 E1 및 E2는 https://jaspar.genereg.net/에 의해 예측되었습니다. SOX2 및 CCAT1 결합 부위 E1을 포함하는 CCAT1 프로모터 서열을 합성하고, CCAT1 3'UTR WT(E1-WT) 및 CCAT1 3'UTR MUT(E1-MUT) 벡터를 형성하였다. 벡터를 pmirGLO(Beyotime)에 클로닝했습니다. 그 후, CCAT1-WT/pmirGLO 또는 CCAT1-MUT/pmirGLO를 sh-SOX2 또는 sh-SOX2 NC로 CD44
+
로 공동 형질감염시켰습니다. HeLa 세포는 각각 2일 동안 용해된 다음 용해됩니다. 루시퍼라제 활성은 루시퍼라제 검출 시스템(Takara, Dalian, China)에 의해 테스트되었습니다.
생물정보학 웹사이트는 CCAT1과 miR-185-3p의 결합 부위 예측 및 분석에 활용되었습니다. CCAT1 및 miR-185-3p의 결합 부위는 이중 루시페라제 리포터 유전자 분석에 의해 확인되었다. miR-185-3p 결합 부위를 포함하는 CCAT1 3'UTR을 구성하였다. CCAT1 3'UTR WT 및 CCAT1 3'UTR MUT를 구성하고 CD44
+
에 대한 miR-185-3p 모방체 및 모방체 NC로 공동 형질감염시켰습니다. 2일 동안 HeLa 세포. 그런 다음, 세포를 용해시키고 루시페라제 검출 시스템(Takara)에 의해 루시퍼라제 활성을 시험하였다. miR-185-3p와 FOXP3의 타겟팅 관계를 확인하기 위해 동일한 방법을 적용했습니다.
통계 분석
모든 데이터는 SPSS 21.0 소프트웨어(IBM Corp. Armonk, NY, USA)로 평가되었습니다. 측정 데이터는 평균 ± 표준편차로 표시하였다. t 테스트는 두 그룹 간의 불일치에 적용되었고 일원 분산 분석(ANOVA)에 이어 그룹 간의 불일치에 대한 Tukey의 다중 비교 테스트가 적용되었습니다. 분류변수는 Fisher의 정확검정으로 평가하였다. p 0.05 미만의 값은 유의한 것으로 간주되었습니다.
<섹션 데이터-제목="결과">
결과
miR-185-3p 발현은 감소하는 반면, SOX2, CCAT1 및 FOXP3 발현은 CC 조직에서 증가하고 SOX2 및 CCAT1 발현은 종양 크기, 림프절과 연결됨 전이(LNM) 및 FIGO 병기
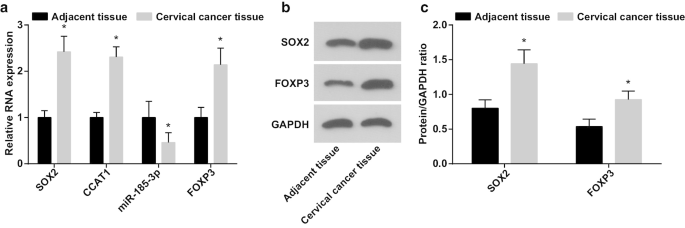
SOX2/CCAT1/miR-185-3p/FOXP3 축이 CC 줄기세포의 증식 및 자가 재생 능력에 미치는 역할을 검출한 결과, CC 조직 및 인접 정상 조직에서 miR-185-3p, SOX2, CCAT1 및 FOXP3 발현이 나타났다. RT-qPCR 및 웨스턴 블롯 분석으로 테스트했습니다. (그림 1a-c) miR-185-3p 발현은 감소한 반면 SOX2, CCAT1 및 FOXP3 발현은 CC 조직에서 증가하는 것으로 나타났습니다.
<그림>
miR-185-3p 발현은 감소하는 반면 SOX2, CCAT1 및 FOXP3 발현은 CC 조직에서 증가합니다. 아 CC 및 인접 정상 조직에서 SOX2 mRNA, CCAT1, miR-185-3p 및 FOXP3 mRNA 발현의 비교. ㄴ CC 및 인접 정상 조직에서 SOX2 및 FOXP3 단백질 발현의 단백질 밴드. ㄷ CC 및 인접 정상 조직에서 SOX2 및 FOXP3 단백질 발현 비교. *p <0.05 대 인접 정상 조직. 측정 데이터는 평균 ± 표준편차로 표시하였고, 두 그룹 간의 비교는 t검정으로 평가
SOX2/CCAT1 발현과 CC의 임상병리학적 특징 사이의 관계를 분석하였다(표 2). CC에서 과발현된 SOX2 및 CCAT1은 종양 크기, LNM 및 FIGO 병기와 연결되어, SOX2 및 CCAT1 발현이 CC 환자의 더 큰 종양 크기, LNM 및 진행된 FIGO 병기를 가진 환자에서 더 높음을 나타냅니다.
miR-185-3p 발현은 감소하는 반면 CD44에서는 SOX2, CCAT1 및 FOXP3 발현 증가
+
헬라 세포
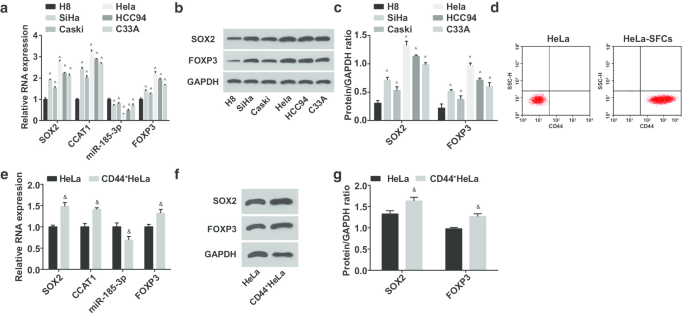
다음으로, 인간 자궁 경부 상피 불멸화 세포 H8 및 CC 세포주 SiHa, CasKi, HeLa, HCC94 및 C33A에서 miR-185-3p, SOX2, CCAT1 및 FOXP3 발현을 테스트했습니다. (그림 2a-c) miR-185-3p 발현은 저하된 반면 SOX2, CCAT1 및 FOXP3는 CC 세포주에서 상승한 것으로 제안되었습니다. 그 중 HeLa 세포의 SOX2/CCAT1/miR-185-3p/FOXP3가 H8 세포와 가장 큰 발현 차이를 보였다. 따라서 HeLa 세포를 선별하고 분류했습니다. HeLa-SFCs는 세포 현탁액 배양에 의해 얻었고, 분류 전후에 세포 표면에서 CD44 발현은 유세포 분석에 의해 확인되었습니다. 결과는 HeLa-SFC에서 CD44의 양성 비율이 분류 전보다 현저히 높아 CC 줄기 세포의 성공적인 분류를 시사함을 강조했습니다(그림 2d). 분류된 줄기세포는 CD44
+
로 명명되었습니다. 헬라 세포. 그런 다음 miR-185-3p, SOX2, CCAT1 및 FOXP3 발현을 HeLa 및 CD44
+
에서 테스트했습니다. HeLa 세포(그림 2e–g). SOX2, CCAT1 및 FOXP3 발현은 상향 조절되고 miR-185-3p는 CD44
+
에서 하향 조절되는 것으로 나타났습니다. 헬라 세포.
<사진>
miR-185-3p 발현은 감소하고 SOX2, CCAT1 및 FOXP3 발현은 CD44에서 증가합니다
+
헬라 세포. 아 H8 및 CC 세포주에서 SOX2 mRNA, CCAT1, miR-185-3p 및 FOXP3 mRNA 발현의 비교. ㄴ H8 및 CC 세포주에서 SOX2 및 FOXP3 단백질 발현의 단백질 밴드. ㄷ H8 및 CC 세포주에서 SOX2 및 FOXP3 단백질 발현의 비교. d 유세포 분석에 의한 분류 전후의 HeLa 세포에서 CD44 발현율의 검출. 이 HeLa 세포와 CD44
+
간의 SOX2 mRNA, CCAT1 및 miR-185-3p 발현 비교 헬라 세포. 에 HeLa 세포 및 CD44
+
에서 SOX2 단백질 발현의 단백질 밴드 헬라 세포. 지 HeLa 세포와 CD44
+
간의 SOX2 단백질 발현 비교 헬라 세포. ^p <0.05 대 H8 세포.
&피 <0.05 vs. HeLa 세포. 측정 데이터는 평균 ± 표준편차로 표시하였고, 두 그룹 간의 비교는 t 검정으로, 여러 그룹 간의 비교는 일원 분산 분석(one-way ANOVA) 후 Tukey 사후 검정으로 평가
SOX2 하향 조절 및 CCAT1 하향 조절은 CD44의 증식, 이동, 침입 및 구체 형성 속도를 감소시키고 세포자멸사를 증가시킵니다
+
헬라 세포
그 후, SOX2 및 CCAT1은 CD44
+
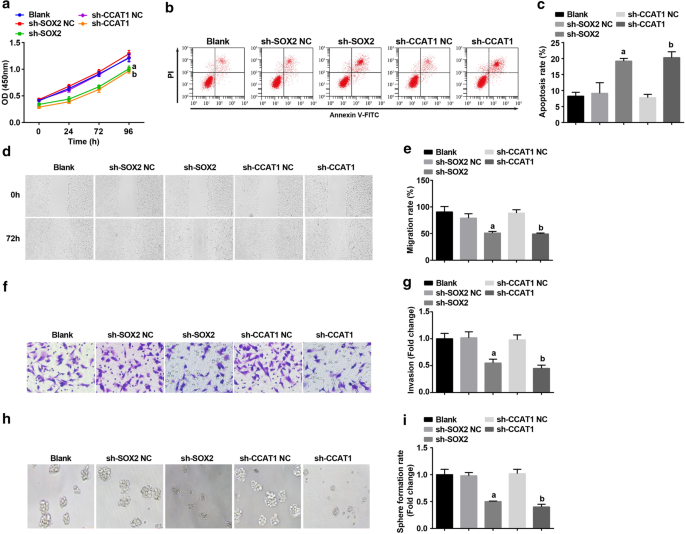
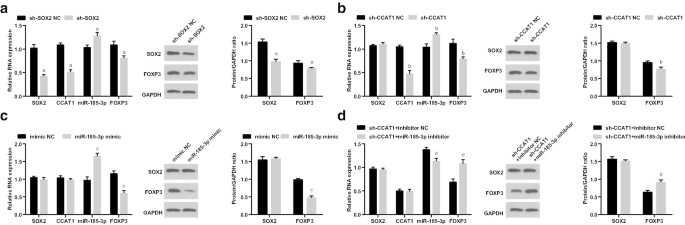
에서 침묵되었습니다. HeLa 세포는 CC 줄기 세포의 증식 및 자가 재생에 미치는 영향을 조사합니다. CCK-8 assay, flow cytometry, scratch test, Transwell assay 및 sphere-forming 실험에 의해 검출, CD44의 증식, 이동, 침습 및 sphere 형성 속도가 억제되었고 세포자멸사 HeLa 세포는 SOX2 및 CCAT1의 억제에 의해 촉진되었습니다(그림 3a-i). SOX2 또는 CCAT1을 억제하면 CC 줄기 세포의 증식 및 자가 재생이 억제됨을 알 수 있습니다.
<그림>
SOX2를 하향 조절하고 CCAT1을 하향 조절하여 증식, 이동, 침습 및 구체 형성 속도를 감소시키고 CD44
+
의 세포자멸사를 증가시킵니다. 헬라 세포. 아 CCK-8 분석은 sh-CCAT1 또는 sh-SOX2로 처리된 세포에서 세포 성장 곡선을 테스트했습니다. ㄴ 유세포 분석은 sh-CCAT1 또는 sh-SOX2로 처리된 세포에서 세포 사멸을 감지했습니다. ㄷ sh-CCAT1 또는 sh-SOX2로 처리한 세포의 세포 사멸 속도 비교. d sh-CCAT1 또는 sh-SOX2로 처리된 세포의 세포 이동은 스크래치 테스트로 테스트되었습니다. 이 sh-CCAT1 또는 sh-SOX2로 처리된 세포에서 세포 이동의 비교. 에 Transwell assay에 의한 sh-CCAT1 또는 sh-SOX2로 처리된 세포에서 세포의 침입 능력 검출. 지 sh-CCAT1 또는 sh-SOX2를 처리한 세포의 침입 능력 비교. 아 구체 형성 실험은 sh-CCAT1 또는 sh-SOX2로 처리된 세포에서 자가 재생 능력을 테스트했습니다. 나 sh-CCAT1 또는 sh-SOX2로 처리된 세포에서 구체 형성 속도 비교. 아피 <0.05 대 sh-SOX2 NC 그룹. 나피 <0.05 대 sh-CCAT1 NC 그룹. 측정 데이터는 평균 ± 표준편차로 표시되었으며, 여러 그룹 간의 비교는 일원 분산 분석(one-way ANOVA)에 이어 Tukey의 사후 검정으로 평가되었습니다.
고갈된 miR-185-3p는 CD44에서 CCAT1의 하향 조절 역할을 역전시킵니다
+
헬라 세포
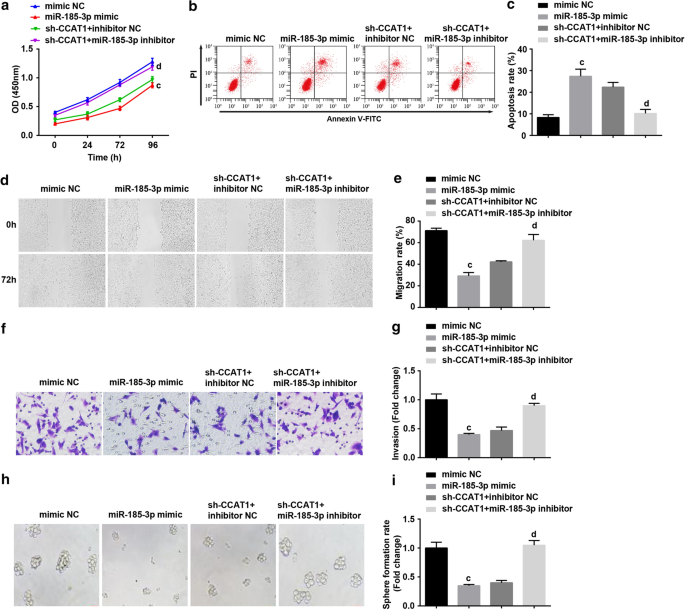
그런 다음 우리는 miR-185-3p가 CC 줄기 세포의 증식과 자기 재생을 조절하는 CCAT1 과정에 관여하는지 조사했습니다. CD44
+
HeLa는 miR-185-3p 모방체로 형질감염되거나 sh-CCAT1 및 miR-185-3p 억제제로 공동 형질감염되었습니다. 결과는 miR-185-3p의 상향 조절이 CD44
+
의 세포 사멸 속도를 높이는 동안 증식, 이동, 침입 및 구체 형성 속도를 현저하게 감소시키는 것으로 나타났습니다. 헬라 세포. miR-185-3p 억제제로 처리된 세포는 CD44
+
의 증식, 이동, 침입, 세포자멸사 및 세포구 형성에서 하향 조절된 CCAT1의 역할을 역전시킬 수 있습니다. HeLa 세포(그림 4a–i).
<그림>
과발현된 miR-185-3p는 증식, 이동, 침입 및 구체 형성 속도를 억제하고 CD44
+
의 세포자멸사를 증가시킵니다. 헬라 세포. 아 CCK-8 분석은 miR-185-3p 모방체로 처리된 세포에서 세포 성장 곡선을 테스트했습니다. ㄴ 유세포 분석은 miR-185-3p 모방으로 처리된 세포에서 세포 사멸을 감지했습니다. ㄷ miR-185-3p 모방체로 처리된 세포의 세포 사멸 속도 비교. d miR-185-3p 모방으로 처리된 세포의 세포 이동은 스크래치 테스트로 테스트되었습니다. 이 miR-185-3p 모방체로 처리된 세포에서 세포 이동의 비교. 에 Transwell 분석에 의한 miR-185-3p 모방체로 처리된 세포의 침입 능력 검출. 지 miR-185-3p 모방체로 처리된 세포의 침입 능력 비교. 아 구체 형성 실험은 miR-185-3p 모방물로 처리된 세포에서 자가 재생 능력을 테스트했습니다. 나 miR-185-3p 모방체로 처리된 세포에서 구체 형성 속도 비교. p <0.05 대 sh-SOX2 NC 그룹. ㄴ 피 <0.05 대 sh-CCAT1 NC 그룹. c p <0.05 대 모방 NC 그룹. d 피 <0.05 대 sh-CCAT1 + 억제제 NC 그룹. 측정 데이터는 평균 ± 표준편차로 표시되었으며, 여러 그룹 간의 비교는 일원 분산 분석(one-way ANOVA)에 이어 Tukey의 사후 검정으로 평가되었습니다.
저발현 SOX2 및 저발현 CCAT1 CD44에서 FOXP3 발현 감소 및 miR-185-3p 발현 증가
+
헬라 세포
그 후 CD44
+
에서 SOX2/CCAT1/miR-185-3p/FOXP3 발현을 조사했습니다. sh-SOX2, sh-CCAT1, miR-185-3p의 형질감염 후 HeLa 세포는 sh-CCAT1 및 miR-185-3p 억제제의 모방 및 공동 형질감염. SOX2, CCAT1 및 FOXP3 발현은 감소한 반면, miR-185-3p 발현은 sh-SOX2로 처리된 세포에서 증가하였다. sh-CCAT1을 처리한 세포에서 CCAT1과 FOXP3의 발현이 감소하였고 miR-185-3p의 발현이 증가하였다. miR-185-3p 모방체가 도입된 세포에서 miR-185-3p 발현이 증가하고 FOXP3 발현이 감소하였다. FOXP3 발현은 증가했고 miR-185-3p 발현은 sh-CCAT1 및 miR-185-3p 억제제로 연속적으로 형질감염된 세포에서 감소했습니다(그림 5a-d).
<그림>
저발현 SOX2 및 저발현 CCAT1은 CD44에서 FOXP3 발현을 감소시키고 miR-185-3p 발현을 증가시킵니다
+
헬라 세포. 아 sh-SOX2 NC 및 sh-SOX2 그룹에서 SOX2/CCAT1/miR-185-3p/FOXP3 발현. ㄴ sh-CCAT1 NC 및 sh-CCAT1 그룹에서 SOX2/CCAT1/miR-185-3p/FOXP3 발현. ㄷ 모방 NC 및 miR-185-3p 모방 그룹에서 SOX2/CCAT1/miR-185-3p/FOXP3 발현. d sh-CCAT1 + 억제제 NC 및 sh-CCAT1 + miR-185-3p 억제제 그룹에서 SOX2/CCAT1/miR-185-3p/FOXP3 발현. p <0.05 대 sh-SOX2 NC 그룹. ㄴ 피 <0.05 대 sh-CCAT1 NC 그룹. c p <0.05 대 모방 NC 그룹. d 피 <0.05 대 sh-CCAT1 + 억제제 NC 그룹. 측정 데이터는 평균 ± 표준편차로 표시되었으며, 여러 그룹 간의 비교는 일원 분산 분석(one-way ANOVA)에 이어 Tukey의 사후 검정으로 평가되었습니다.
SOX2는 miR-185-3p 발현에 영향을 미치는 CCAT1에 결합하고 FOXP3는 miR-185-3p의 표적 유전자입니다.
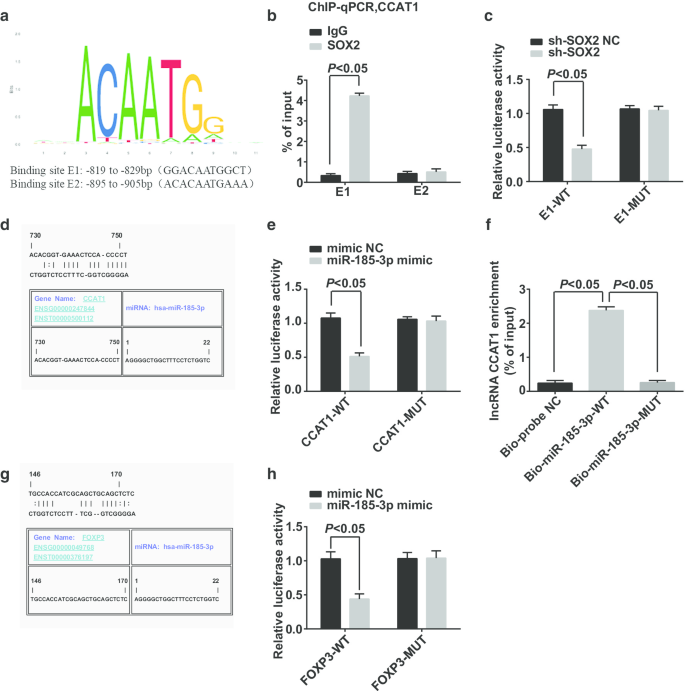
CCAT1 프로모터 영역 전사 인자의 잠재적 결합 부위는 https://jaspar.genereg.net/웹사이트에서 예측 및 분석되었으며, SOX2 및 CCAT1이 CCAT1 프로모터 영역에 잠재적인 결합 부위를 갖는 것으로 나타났습니다(그림 6a). ChIP-qPCR은 다음과 같이 보고했습니다(그림 6b):IgG 그룹과 달리 E1 결합 부위에서 SOX2 그룹에서 더 많은 CCAT1 프로모터 단편이 농축되어 SOX2가 E1 부위에서 CCAT1 프로모터에 결합되었고 SOX2가 CCAT1의 규제에 관여합니다. 이중 루시페라제 리포터 유전자 분석 결과(그림 6c):sh-SOX2 및 E1-WT로 동시 형질감염된 세포에서 루시퍼라제 활성이 억제되었으며, 이는 SOX2가 CCAT1에 결합할 수 있음을 나타냅니다.
<그림>
SOX2는 miR-185-3p 발현에 영향을 미치는 CCAT1에 결합할 수 있으며, FOXP3는 miR-185-3p의 표적 유전자이다. 아 생물정보학 사이트에 의한 SOX2 및 CCAT1 프로모터 영역의 결합 사이트 예측. ㄴ ChIP-qPCR 실험은 SOX2와 CCAT1 간의 결합 관계를 확인했습니다. ㄷ 이중 루시퍼라제 리포터 유전자 분석에 의해 확인된 SOX2 및 CCAT1의 결합 부위. d 생물정보학 사이트에 의한 CCAT1 및 miR-185-3p의 결합 사이트 예측. 이 이중 루시퍼라제 리포터 유전자 분석에 의한 CCAT1 및 miR-185-3p의 결합 확인. 에 RNA 풀다운 분석으로 확인된 세포에서 CCAT1과 miR-185-3p 간의 결합 관계. 지 생물정보학 웹사이트에서 miR-185-3p와 FOXP3 간의 표적화 관계 예측. 아 이중 루시퍼라제 리포터 유전자 분석에 의한 miR-185-3p와 FOXP3 간의 표적화 관계 식별. 측정 데이터는 평균 ± 표준편차로 표시하였고, 두 그룹 간의 비교는 t검정으로 평가
Jefferson 웹사이트는 CCAT1이 miR-185-3p에 결합할 수 있다고 예측했습니다(그림 6d). 이중 루시퍼라제 리포터 유전자 분석은 (그림 6e) miR-185-3p 모방체와 CCAT1-WT가 도입된 세포에서 루시페라제 활성이 감소했다고 보고했으며, 이는 miR-185-3p가 CCAT1에 결합할 수 있음을 시사합니다. RNA 풀다운 분석을 사용하여 CCAT1이 miR-185-3p와 결합할 수 있는지 여부를 확인했습니다. 결과는 (그림 6f) Bio-miR-185-3p-WT를 처리한 세포에서 CCAT1의 농축 수준이 크게 증가한 반면, Bio-miR-185-3p-MUT를 처리한 세포에서 CCAT1의 농축 수준은 큰 차이가 없습니다. 이 결과는 CCAT1이 miR-185-3p를 흡착하여 miR-185-3p 발현에 영향을 줄 수 있음을 보여주었습니다.
miR-185-3p와 FOXP3 사이의 목표 관계는 Jefferson 웹사이트에서 예측했습니다(그림 6g). 이중 루시퍼라제 리포터 유전자 분석은 (그림 6h) FOXP3-WT 및 miR-185-3p 모방체가 CD44
+
로 공동 형질감염된 후 세포의 상대적 루시페라제 활성이 극적으로 감소되었음을 확인했습니다. HeLa 세포에 비해 miR-185-3p 모방체로 공동 형질감염된 FOXP3-MUT는 세포의 상대적인 루시페라제 활성에 영향을 미치지 않아 miR-185-3p가 FOXP3를 표적으로 함을 시사합니다.
토론
CC는 유방암, 결장암, 폐암에 이어 세계에서 네 번째로 흔한 여성의 악성종양입니다[3]. SOX2 발현이 양성인 CC 세포는 암 줄기세포의 특성을 보인다고 보고되었다[21]. 한 연구에 따르면 CCAT1은 CC와 관련된 필수 발암성 lncRNA이며 CC 세포 성장 및 침입을 촉진하는 역할을 한다고 보고되었습니다[11]. 또 다른 연구에서는 miR-185-3p가 비인두암의 방사선 감수성을 예측하고 암세포의 성장과 세포 사멸을 조절할 수 있음을 밝혔습니다[22]. FOXP3에 대한 순환 자가항체는 자궁경부 병변의 지속적인 진행을 반영하며 CC의 조기 예후를 위한 잠재적 바이오마커가 될 수 있다고 보고되었습니다[23]. 현재 연구는 SOX2/CCAT1/miR-185-3p/FOXP3 축이 CC 줄기 세포의 증식 및 자가 재생 능력에 어떤 영향을 미치는지 알아보기 위해 고안되었습니다.
우리의 발견에 기초하여, SOX2 및 CCAT1 발현은 종양 크기, LNM 및 진행된 FIGO와 관련된 CC 조직 및 세포에서 증가했습니다. Functionally proved, down-regulating SOX2 and CCAT1 declined proliferation, migration, invasion and sphere cells number and increased apoptosis of CC stem cells. Similar to our study, SOX2 expression trends to increase in CC [24, 25]. Moreover, SOX2 expression is also up-regulated in CC cells derived from cancer stem cells [26]. Overexpressed SOX2 was suggested to link with clinicopathological characteristics of patients with several types of cancer, not limited to CC. For example, it was suggested that up-regulated SOX2 shows in cervical squamous cell carcinoma patients staged in FIGO I-II [27]. Moreover, SOX2 expression is linked to LNM in oral squamous cell carcinoma [28]. When it comes to the molecular function of SOX2 for cancer progression, there is an observational work presenting that down-regulated SOX-2 suppresses cell migration and invasion of cervical squamous cell carcinoma [29]. Meanwhile, another research has offered a proof that up-regulated SOX2 enhances CC cell clonogenicity, proliferation and tumorigenicity in vitro and in vivo than control cells [30].
Concerning to the regulatory relation between SOX2 and CCAT1, an existed study has presented that silencing SOX2 markedly reduces CCAT1 mRNA level [31]. As to the role of CCAT1 in cancers, a study has showed that CCAT1 expression is markedly elevated in CC tissues versus in the adjacent normal tissues [11, 12]. Of note, CCAT1 overexpression in CC is positively related to the tumor size [12]. In terms of the role of CCAT1 in cancer cell activity, there is a research highlighting that overexpressed CCAT1 accelerates CC cell proliferation, colony formation and invasion [11]. Interestingly, a previous research has demonstrated that the cell viability, invasive and migratory abilities are declined via knocking down CCAT1 [12]. Anyway, the functional effect of SOX2 and CCAT1 in other cancers was similar to that in CC.
Afterward, our research revealed that CCAT1 could bind to miR-185-3p, the down-regulated CCAT1 in CC and overexpressing miR-185-3p suppressed the proliferation and self-renewal abilities of CC stem cells. It is reported that CCAT1 and miR-185-3p are negatively correlated [13]. Furthermore, a result reported that a reduction is seen in miR-185-3p expression in radioresistant nasopharyngeal carcinoma cases [22]. Regarding to the suppressive function of miR-185-3p in cancer cell aggressiveness, a study has revealed that up-regulation of miR-185-3p suppresses the invasive and metastatic properties of nasopharyngeal carcinoma cells [32]. Furthermore, Zou et al. have suggested that restored miR-185 represses breast cancer cell growth and invasion [33]. There is a article finding that up-regulation of miR-185 declines the proliferation, invasion and colony formation capacities of non-small cell lung cancer cells in vitro [34]. It is presented that in vitro cell proliferation, invasion and migration as well as in vivo tumor growth are suppressed via miR-185-overexpressing in non-small cell lung cancer cells [35]. From those studies, the anti-tumor role of miR-185-3p in the present study was consistent with previous researches.
To proceed, we unveiled that miR-185-3p targeted FOXP3, the overexpressed gene in CC to regulate CC stem cell activities. In fact, FOXP3, the regulator of SOX2 cancer stem-like cell marker in colon cancer [36], has been investigated in CC, showing an up-regulation in CC cells [19] [37]. It was evidenced that elevating FOXP3 promotes the formation of tumor spheres and stimulates the stemness of non-small cell lung cancer cells [38].
Conclusion
Collectively, we explored for the first time that SOX2 transcription could activate CCAT1, thereby inhibiting miR-185-3p and regulating FOXP3 to promote the proliferation and self-renewal of CC stem cells, which is a potential avenue to treat CC. Additionally, however, limitations in this present study still exist in the relatively small trial size in the designed experiment. Thus, clinical researches might be further carried out to detect the efficacy for the treatment of CC.