골수 중간엽 줄기 세포 유래 엑소좀 MicroRNA-133a는 MAML1 억제를 통해 바이러스성 심근염 쥐에서 심근 섬유증 및 상피-중간엽 전환을 억제합니다.
초록
심근염은 효과적인 치료 없이 심근의 국소화 또는 확산 염증을 특징으로 하는 질병이다. 이 연구는 조절을 통해 바이러스성 심근염(VMC) 쥐의 심근 섬유증 및 상피-중간엽 전이(EMT)에 대한 골수 중간엽 줄기세포 유래 엑소좀(BMSC-Exo)에서 분비되는 microRNA-133(miR-133)의 조절 메커니즘을 탐구했습니다. 마스터마인드 1(MAML1). 쥐의 BMSC를 분리 배양하여 면역 표현형과 골 형성 및 지방 형성 능력을 확인하고 BMSC-Exo를 추출하여 확인했습니다. 초원심분리를 통해 엑소좀을 얻었고, 투과전자현미경과 웨스턴 블롯 분석을 통해 확인하였다. VMC 모델 제작을 위해 Rat에 Coxsackie B3 바이러스를 주입하고, 동물 실험(NC
Exo
)과 동일한 방법으로 심근세포를 분리, 배양 및 그룹화 , Ad-miR-133a
엑소
, Adas-miR-133a
엑소
). 염증, 세포 사멸, EMT, 섬유증 및 세포 생존에서 엑소좀 miR-133a 및 MAML1의 역할을 파악하기 위해 생체 내 및 시험관 내 실험을 수행했습니다. miR-133a와 MAML1 간의 표적화 관계는 이중 루시페라제 리포터 유전자 분석에 의해 확인되었습니다. BMSC-Exo는 VMC 쥐에서 miR-133a 발현을 증가시켰고 VMC 쥐의 심장 기능과 심근 섬유증을 효과적으로 개선하고 심근세포 생존력을 증가시켰으며 EMT 과정을 억제했습니다. 엑소좀에서 증가된 miR-133a는 개선을 강화했습니다. 침묵의 miR-133a는 VMC 쥐에 대한 BMSC-Exo의 효과를 효과적으로 역전시켰습니다. miR-133a는 MAML1을 표적으로 했습니다. MAML1의 억제는 VMC 쥐의 심장 기능과 심근 섬유증을 개선하고 VMC 쥐에 대한 miR-133a 침묵 엑소좀의 효과를 역전시킬 수 있습니다. 우리의 연구는 상승된 엑소좀 miR-133a가 MAML1을 하향 조절함으로써 VMC가 있는 쥐에서 심근 섬유증과 EMT를 억제하여 심근염의 진행을 억제함을 시사합니다.
소개
심근염은 심장 근육 세포의 염증성 질환으로 간주됩니다[1]. 심근염은 분명히 여성에 비해 남성에게 더 친숙합니다[2]. 바이러스성 심근염(VMC)은 젊은 사람들에서 확장성 심근병증(DCM)과 급사를 일으키는 주요 요인입니다[3]. 심근염의 임상적 양상은 모호한 징후와 증상을 보이는 무증상 상태에서부터 심인성 쇼크 및 부정맥으로 고통받는 면역 세포 및 바이러스에 의한 심각한 심근 파괴에 이르기까지 다양하다[1]. 심근염은 비감염성 유도제와 함께 바이러스, 박테리아, 클라미디아, 리케차, 진균 및 원생동물로 구성된 다양한 감염 요소에 의해 유도될 수 있습니다. 그 중 바이러스 감염이 가장 흔한 원인이며, 특히 소아에서 발생합니다[4]. 심근염을 일으키는 가장 중요한 바이러스인 Coxsackie B3 바이러스(CVB3)는 VMC의 발병기전에 산화 스트레스 반응과 세포자멸사를 유발할 수 있지만 VMC에 대한 특별한 치료는 아직 보고되지 않았다[5]. 더욱이 VMC의 발병기전은 잘 알려져 있지 않으며 정확한 임상적 치료도 부족하다[3]. 따라서 질병의 예후를 개선하기 위해서는 새로운 표적이 시급합니다.
MicroRNA(miRNA)는 단백질 코딩 유전자의 발현을 조절할 수 있는 내인성 비암호화 RNA입니다[6]. MiR-133a는 심장 특이적 miRNA 중 하나로 심근 경색증(MI)을 포함하는 심장 발달 및 일부 심혈관 질환과 관련이 있습니다[7]. 또한 miR-133a는 만성 샤가스병 심근병증에서 비정상적으로 발현되는 것이다[8]. 또한 심근의 miR-133a 수치는 염증성 심근병증의 염증, 좌심실 기능 및 임상 결과와 관련이 있습니다[9]. MiRNA는 마우스 및 인간 비만 세포에서 유래한 엑소좀에서 발견되었습니다[10]. 엑소좀, 대부분의 세포 종류에서 방출되는 나노 크기의 소포는 다양한 생물학적 유체에서 발견됩니다[11]. 엑소좀은 자신의 화물을 수용 세포로 전달할 수 있으며, 이는 수용 세포의 생화학적 구성과 신호 전달 경로를 변화시키는 것으로 나타났습니다[12, 13]. 변화된 엑소좀 miRNA가 CVB3 유발 심근염의 발병기전과 관련이 있다는 증거가 있습니다[14]. 저산소 상태로 조절된 골수 중간엽 줄기 세포의 엑소좀 miR-125b-5p는 심근세포의 세포자멸사를 감소시키고 허혈성 심장 복구를 향상시킨다[15]. 또한 MSC의 엑소좀 miR-25-3p는 심근세포의 세포자멸사 및 염증 반응을 감소시켜 MI를 완화합니다[16]. 흥미롭게도 엑소좀 심장 발현 miR-133a는 심장 트로포닌 심장 트로포닌-I에 연결되어 있습니다[17]. Mastermind-like 1(MAML1)은 심근 허혈/재관류(I/R) 손상과 관련된 것으로 보고된 우리 연구에서 miR-133a의 교차 선별된 다운스트림 유전자였습니다[18]. 또한 최근 연구에서는 MAML1 녹다운이 간 섬유화증에서 항섬유화 기능을 갖는다고 언급했습니다[19].
이전 연구에 의해 밝혀진 바에 따르면 BMSC 유래 엑소좀 miR-133a가 심근염을 매개할 수 있는지 여부가 궁금합니다. 따라서 이 연구는 BMSC 유래 엑소좀(BMSC-Exo)에 의해 셔틀된 miR-133이 MAML1의 조절을 통해 VMC 쥐의 심근 섬유증 및 상피-중간엽 전이(EMT)를 개선한다는 가설에서 시작되었습니다.
자료 및 방법
윤리적 승인
연구는 저장대학교 의과대학 제4부속병원의 기관 동물 관리 및 사용 위원회의 허가를 받았습니다. 동물을 인도적으로 대했습니다.
BMSC의 분리
실험 동물은 성숙한 특이적 병원균이 없는(SPF) 등급 Sprague-Dawley(SD) 수컷 쥐였습니다(Zhejiang University School of Medicine, Zhejiang, China의 실험 동물 센터). 펜토바르비탈 나트륨을 복강내 주사하여 쥐를 안락사시키고 75% 알코올로 살균하였다. 대퇴골과 경골을 깨끗한 테이블에서 꺼내고 근육과 결합 조직을 제거하고 골수강을 저포도당 Dulbecco의 Modified Eagle Medium(DMEM)으로 반복해서 헹구었습니다. 액체를 원심분리하여 침전물을 회수하고 재현탁하고 24시간 동안 배양했습니다(배지는 2-3일마다 교체했습니다). 대수 단계로 성장할 때 BMSC를 0.25% 트립신(Gibco, Carlsbad, California, USA)으로 분리하고, 원심분리하고 MSC 배양 용액(Cyagen Biosciences Inc., Guangzhou, China)에 재현탁했습니다. 현탁액을 1:2의 비율로 통과시켰다. 위의 조작을 반복하였고, 4번째 계대 BMSC를 후속 실험에 사용하였다.
BMSC 식별
대수 성장에서 4차 계대 BMSC의 표면 항원은 유세포 분석에 의해 확인되었습니다. BMSC를 에틸렌 디아민 테트라아세트산을 함유하는 0.25% 트립신(1mL)으로 분리하고, 원심분리하고, 적절한 인산완충식염수(PBS)로 재현탁하고 151g에서 원심분리했습니다. 그런 다음 BMSC를 2% 신선한 소 태아 혈청(FBS)(Gibco)을 함유하는 PBS로 재현탁하여 단일 세포 현탁액을 제조하였다. FITC-CD34, PE-CD29 및 PE-CD44 단일클론 항체(각각 5μL, BD Biosciences, Franklin Lakes, NJ, USA)를 세포 현탁액(100μL)과 함께 인큐베이션하고, 151g에서 원심분리하고, 500g으로 재현탁시켰다. 1% 파라포름알데히드를 함유하고 30분 동안 고정된 μL PBS. 배경 마커는 호모타입 대조군의 모노클로날 항체를 사용하여 식별되었습니다.
유세포 분석:단일 세포 현탁액을 고정하고 151g에서 원심분리했습니다. 그런 다음 BMSC를 1% 파라포름알데히드를 포함하는 PBS로 재현탁하고 MACS Quart 유세포 분석기에서 테스트하고 해당 소프트웨어로 분석했습니다.
BMSC의 골형성 및 지방형성 유도
4차 계대에서 BMSC를 200개 세포/mL로 6개의 웰 플레이트에 접종했습니다. 조골 세포 유도 용액과 지방 생성 유도 용액(Cyagen Biosciences Inc.)을 60-70% 합류의 BMSC에 첨가했습니다. 다른 두 웰의 BMSC에는 유도 액체가 대조군으로 추가되지 않았습니다. BMSC를 14일 동안 유도하고 4% 파라포름알데히드로 고정시켰다. 그런 다음 분화된 조골세포와 지방세포를 Alizarin red 염색과 oil red O 염색(Wuhan Pulande Biological Technology Co., Ltd., Wuhan, China)으로 시행하고 현미경으로 관찰하였다.
엑소솜 분리 및 식별
4차 계대에서 BMSC를 48시간 동안 배양하여 상층액을 수집한 다음 원심분리(800g 및 2000g)하고 0.22μm 및 100,000MW 필터 멤브레인으로 여과하고 원심분리(100,000g)하여 침전물을 수집했습니다. 그 후, 침전물을 PBS로 재현탁하고, 다시 100,00g에서 원심분리하여 엑소좀 침전물을 얻었다. PBS 중의 BMSC-Exo 현탁액은 웨스턴 블롯 분석(Proteintech, Chicago, IL, USA)에 의한 비신코닌산(bicinchoninic acid, BCA) 및 엑소좀 제조 단백질 검출(CD63, CD81 및 CD9)에 의한 농도 검출을 거쳤다.
재조합 아데노바이러스 감염은 BMSC의 miR-133a 유전자 변형을 매개합니다.
BMSC를 밤새 계대시켰다. 정상 대조군(동량 PBS), miR-133a 음성 대조군(NC), miR-133a 과발현(Ad-miR-133a), miR-133a 저발현(Adas-miR-133a)( Hanbio Biotechnology Co., Ltd., Shanghai, China)는 100 MOI(multiplicity of infection)에 따라 BMSC로 형질감염되었다. BMSC를 배양하고 상응하는 엑소좀(NC
Exo
, NC
엑소
, Ad-miR-133a
엑소
및 Adas-miR-133a
엑소
) 초원심분리를 통해 얻었다[20].
쥐의 VMC 모델 확립 및 실험 동물 그룹화
성숙한 수컷 SPF 등급 SD 쥐를 각각 8마리씩 10개 그룹으로 나누었습니다. Coxsackievirus B3(CVB3)는 중국 의학 아카데미(중국 베이징)의 의학 생명 공학 연구소에서 제공했습니다.
CVB3(10 mg/kg)는 쥐에게 복강내 주사하고 PBS 또는 BMSC-Exo(100 μg)는 꼬리 정맥을 통해 주사하였다. 대조군에 대한 정상 래트에 CVB3 배양액 및 PBS를 주사하였다. 10 mg/kg CVB3를 주사한 쥐에게 PBS, MSC
exo
를 추가로 주사했습니다. , NC
엑소
, Adas-miR-133a
엑소
, Ad-miR-133a
엑소
, si-NC 또는 si-MAML1(RIBOBIO, 중국 광저우).
쥐에게 7일 동안 지속적으로 주사하여 안구 혈액을 채취하였다. 혈액을 원심분리하고 혈청을 수집하고 하위 포장하여 - 20 °C에서 보관했습니다. 쥐를 안락사시킨 후 심장 표본을 채취하여 10% 포름알데히드로 고정하고 구배 알코올로 탈수하고 크실렌으로 제거하고 파라핀으로 포매한 후 조직학적 관찰을 위해 절편화했습니다. 분자생물학 실험 재료로 섹션의 일부를 - 80 °C에 두었습니다.
심초음파검사
바이러스 주사 후 7일째에 쥐에게 펜토바르비탈 나트륨 25 mg/kg을 복강내 주사하였다. 완전 마취 후 전극침과 연결된 심전도기의 사지 리드를 쥐의 사지 끝에 피하로 삽입하고 사지 리드 심전도를 기록하였다. 그런 다음 쥐를 약간 왼쪽으로 앙와위 자세로 고정하고 가슴을 제모하고 II 심전도를 연결하여 흉골 4 챔버 심장 섹션에서 대동맥 혈류 펄스의 도플러 스펙트럼을 얻었다. 지표에는 좌심실 후벽 두께(LVPW), 좌심실 수축기말 직경(LVID), 좌심실 단축률(FS) 및 좌심실 박출률(LVEF)이 포함됩니다.
헤마톡실린-에오신(HE) 염색
조직을 4% 파라포름알데히드로 고정하고, 탈수하고, 투명하게 하고, 파라핀으로 포매하였다. 그런 다음 4-μm 섹션을 왁스 제거하고 헤마톡실린(Servicebio, 무한, 중국)으로 염색하고 1% 염산 알코올로 분화한 후 파란색으로 되돌린 다음 에오신으로 염색하고 탈수하고 크실렌으로 지우고 중성 검으로 밀봉하고, 광학현미경(Olympus, Tokyo, Japan)으로 관찰했습니다.
마손 콜라겐 염색
파라핀 절편은 왁스를 제거하고 헤마톡실린으로 2분 이내에 염색하고 Lichun 마젠타 용액으로 염색하고 0.5% 빙초산 용액으로 빠르게 헹구었습니다. 그런 다음 절편을 1% 인산알루미늄 수용액으로 염색하고 짙은 빨간색에서 밝은 빨간색, 분홍색으로 염색하여 현미경으로 관찰하였다. 그런 다음 섹션을 Aniline blue(Pulande)로 염색하고, 일반적으로 자일렌으로 탈수하고 밀봉했습니다. Medical Image 분석 소프트웨어 Image-Proplus 6.0을 사용하여 콜라겐 섬유의 양성 염색 면적과 콜라겐 부피 분율(CVF) =콜라겐 면적/총 필드 면적을 측정했습니다. 콜라겐 섬유의 염색 위치와 색상을 구별하였다(심근세포는 적색, 콜라겐 섬유는 청색 띠 또는 세포간 공간의 균질한 구조).
말단 Deoxynucleotidyl Transferase-Mediated Deoxyuridine Triphosphate-Biotin Nick End-Labeling(TUNEL) 염색
파라핀 섹션은 왁스를 제거하고 시트레이트 완충액에 넣고 350W에서 10분 동안 굽습니다. 절편에 50μL TUNEL 용액을 첨가하고, 50μL 전환제-과산화효소와 결합하고, DAB로 현상하고, 현미경으로 관찰하였다. 절편을 헤마톡실린에 넣고 95% 에탄올 I-II에 담그고 무수 에탄올 I-II, 크실렌 I-II와 결합하고 밀봉했습니다. 광학현미경으로 분석한 결과입니다.
효소 결합 면역흡착 분석법(ELISA)
종양 괴사 인자 α(TNF-α), 인터루킨(IL)-1β 및 IL-6은 ELISA 키트(BOSTER Biological Technology Co. Ltd., 무한, 중국)에 의해 검출되었습니다. 안구 혈액을 604g에서 원심분리하여 상부 혈청을 수집하였다. 세포 배양액에서 원심분리하여 얻은 상등액을 세포 실험에서 검출하였다. 샘플의 희석 표준에는 7가지 농도 구배가 있었습니다. 블랭크 웰에 샘플 희석제를 결합하고, 다른 웰에 테트라메틸벤지딘(TMB)을 첨가하고, 각 농도에 대해 2개의 이중 웰을 설정하였다. 샘플 웰을 50 μL의 샘플 희석제 및 샘플과 차례로 결합했습니다. 각 웰은 1시간 동안 100μL의 1차 항체(TMB 웰 제외)와 300μL의 0.01M tris-buffered saline(TBS) 및 100μL Avidin-Biotin-Peroxidase Complex working solution(TMB well 제외)과 반응했습니다. ). 그런 다음 각 웰에 300μL의 0.01M TBS를 첨가하고 100μL TMB와 함께 인큐베이션했습니다. 각 well의 OD(Optical Density) 값과 농도를 바로 측정하여 표준곡선을 그렸습니다.
역전사 정량적 중합효소 연쇄 반응(RT-qPCR)
심근 조직 및 심근세포에서 MiR-133a, 콜라겐 I, 콜라겐 III, α-SMA, TGF-β1, CTGF, E-cadherin 및 FSP-1 발현이 RT-qPCR을 통해 검출되었습니다. 전체 RNA는 심근 세포 또는 심근 조직에서 추출하고 RNA 추출 키트(Takara, Dalian, China)를 통해 cDNA로 역전사하고, RT-PCR 프라이머는 Invitrogen(Guangzhou, China)을 통해 합성했으며, 서열은 표 1에 나와 있습니다. 상대적 정량적 유전자 발현은 2
-△△Ct에 따른 loading control 유전자로 GAPDH(glyceraldehyde-3-phosphate dehydrogenase) 또는 U6을 사용하여 분석하였다.
방법.
서부 얼룩 분석
쥐를 마취로 안락사시켰다. 심근 조직을 동결하고 액체 질소에서 분쇄했습니다. 그 다음, 프로테아제 억제제 페닐메탄설포닐플루오라이드 스톡 용액을 세포 용해 완충액과 1:100의 비율로 혼합하였다(Beyotime Biotechnology Co., Ltd., Shanghai, China). 샘플을 혼합 용액으로 용해하고 세포의 단백질을 추출하였다. 총 단백질 농도는 BCA 키트로 검출되었습니다. 샘플을 4:1로 5 × 로딩 완충액과 혼합하고, 끓는 수조에서 10분 동안 실시하고, 얼음으로 목욕시킨 후 원심분리하였다. 전기영동 분리를 하고 전기이동액을 이용하여 단백질을 폴리비닐리덴 플루오라이드(Servicebio) 막으로 옮겼다. 그런 다음 막을 5% 탈지 분유로 차단하고 1차 항체 CD63, CD81 및 CD9(Proteintech의 토끼 항-쥐 다클론 항체, 1:100), MAML1(ab65090, Abcam, MA, USA, 1:1000) 및 GAPDH(Santa Cruz Biotechnology, Inc, Santa Cruz, CA, USA, 1:1000). 그런 다음 멤브레인에 2차 항체인 양고추냉이 퍼옥시다제 표지된 IgG(Cell Signaling Technology, Beverly, MA, USA, 1:1000)를 적하하고 향상된 화학 발광 반응 용액(Pierce, Rockford, IL, USA)에 담그었습니다. GAPDH를 로딩 컨트롤로 사용하여 ImageJ2x 소프트웨어로 단백질 각인 이미지를 분석했습니다.
심근세포 배양 및 통로
3-5일령의 SD 쥐(중국 저장성 저장대학교 의과대학 실험동물센터)를 채취하였다. 심실 부분을 미리 냉각된 Hank's Balanced Salt Solution으로 헹구고 작은 조각으로 자르고 0.25% 트립신으로 분리했습니다. 조각에 적절한 양의 10% 완전 배지를 첨가하여 분리를 종결하고 151g에서 원심분리하였다. 20% FBS를 함유하는 DMEM을 세포 재현탁에 적용하였다. 차별적 부착법으로 심근세포를 정제하고, 트립판 블루 염색을 통해 생존율을 관찰하고, 생존한 심근세포를 배양하였다. 24시간 후 심근세포가 벽에 달라붙어 맥박이 뛰기 시작했습니다. 72시간 후, pseudopodia가 확장되었습니다.
심근세포의 VMC 모델 구축
대수증식기 4계대 심근세포를 선별하여 MSC
Exo
감염 , NC
엑소
, Adas-miR-133a
엑소
및 Ad-miR-133a
엑소
. 100 Tcid50 CVB3 바이러스 용액(100 μL)을 세포에 첨가하여 세포 VMC 모델을 유도하였다. 동시에 대조용 세포에 동량의 유지 용액을 첨가하고 47시간 배양 동안 1시간 감염 후 상응하는 엑소좀을 심근세포에 결합시켰다.
세포 계수 키트(CCK)-8 분석
CCK-8 세포 검출 키트(Beyotime)를 적용하여 심근세포의 생존율을 검출하였다. 대수기(logarithmic phase)로 성장했을 때, 세포는 0.25% 트립신으로 분리되었고, 2.5 × 10
4
에서 96웰 세포 배양 플레이트에 파종되었습니다. 세포/웰당. CCK-8 용액(10 μL/well)과 결합하여 세포를 1-4시간 동안 지속적으로 배양하고 OD450 nm 값은 마이크로플레이트 리더를 통해 측정되었습니다.
유세포분석
AnnexinV-APC/propidium iodide (PI) 이중 염색 방법을 적용하여 세포 사멸을 검출했습니다. 세포를 원심분리하고 250μL 결합 완충액(4mL 결합 완충액+ 12mL 탈이온수)으로 재현탁하고 1 × 10
6
으로 조정했습니다. 세포/mL. 100μL 세포 현탁액에 5μL Annexin V-APC(BD Biosciences) 및 5μL PI 용액(BD Biosciences)을 첨가하고 유세포 분석기에 로드하고 컴퓨터에 의해 자동으로 분석했습니다.
이중 루시퍼라제 리포터 유전자 분석
MAML1 3-비번역 영역(UTR)의 야생형(wt) 또는 돌연변이형(mut) 서열을 pGL3-M 벡터(Promega, WI, USA)에 클로닝한 다음, MAML1-3-UTR-wt 또는 MAML1- 3-UTR-mut가 생성되었습니다. 벡터는 miR-133a 모방체 또는 NC와 함께 Lipofectamine 2000을 통해 심근세포에 공동 형질감염되었습니다. 루시퍼라제 활성은 이중 루시페라제 리포터 유전자 시스템(Promega)에 의해 48시간 후에 테스트되었습니다. [21]
RNA 면역침전(RIP) 분석
RIP 키트(Millipore, USA)를 사용하여 MAML1과 miR-133a의 결합을 감지했습니다. 세포를 방사선 면역침전 분석 완충액(P0013B, Beyotime, Shanghai, China)에 의해 용해시키고, 1400g에서 원심분리하고, 공침에 대한 항체와 함께 인큐베이션하였다. 자기 비드(50μL)를 100μL RIP 세척 완충액에 재현탁하고 5g 항-MAML1 항체(1g/mL, ab155786) 또는 IgG(1:100, ab172730)와 함께 인큐베이션했습니다. 자기 비드-항체 복합체를 900μL RIP 세척 완충액에 재현탁하고, 100μL 세포 추출물과 상호작용하고, 프로테이나제 K로 분해하고, RT-qPCR로 검출했습니다[22].
통계 분석
데이터 분석에는 SPSS 21.0 통계 소프트웨어(IBM Corp. Armonk, NY, USA)가 적용되었습니다. 측정 데이터는 평균 ± 표준편차로 표현하였다. t 테스트는 두 그룹 간의 비교에 적용되었습니다. 일원 분산 분석(ANOVA)은 그룹 간 비교에 사용되었고 Tukey의 사후 테스트는 쌍별 비교에 사용되었습니다. 예측 변수는 P에서 유의한 경우 유지되었습니다. 0.05 이하의 값입니다.
<섹션 데이터-제목="결과">
결과
BMSC 및 BMSC-Exo 식별
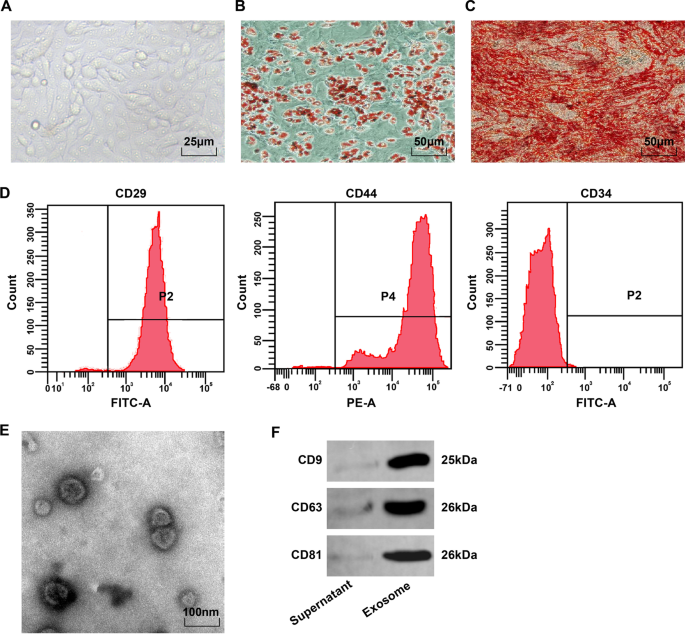
현미경으로 볼 때 BMSC는 방추형이고 둥글고 소용돌이 또는 방사형 패턴으로 벽에 부착되었습니다(그림 1A). 지방 생성 유도의 오일 레드 O 염색 후, 4번째 BMSC의 지질 방울은 적색이었고, 다양한 크기의 둥근 지질 방울이 있었다(도 1B). 골형성 유도 후 석회화 결절을 발현하는 세포는 alizarin red 염색 후 적색을 보였고 석회화 결절과 중첩 세포의 분포가 고르지 않았다(Fig. 1C). 유세포 분석은 MSC 마커 CD29 및 CD44(> 95%)가 발현되었지만 조혈 줄기 세포 표면 항원 CD34(<95%)는 발현되지 않은 것으로 나타났습니다(그림 1D). 이러한 결과는 BMSC가 고순도이고 국제 세포 치료 학회의 MSC 표준을 준수함을 나타냅니다.
<사진>
BMSC 표현형 관찰 및 BMSC-Exo 식별. A 4번째 구절에서 BMSC의 형태학적 관찰; 나 지방세포의 oil red O 염색 결과; ㄷ 조골세포의 알리자린 레드 염색 결과; 디 유세포 분석을 통해 검출된 BMSC의 표현형. 이 BMSC-Exo의 전자현미경 관찰; F CD9, CD63 및 CD81의 단백질 밴드
투과전자현미경은 BMSC-Exos가 명확한 주변막 구조, 다양한 크기 및 40-100 nm의 직경을 갖는 타원형 소포임을 관찰하였다(그림 1E). Western blot 분석 결과, 추출된 생성물이 CD9, CD63, CD81의 특징적인 단백질 유래 엑소좀을 발현함을 확인하였다(Fig. 1F).
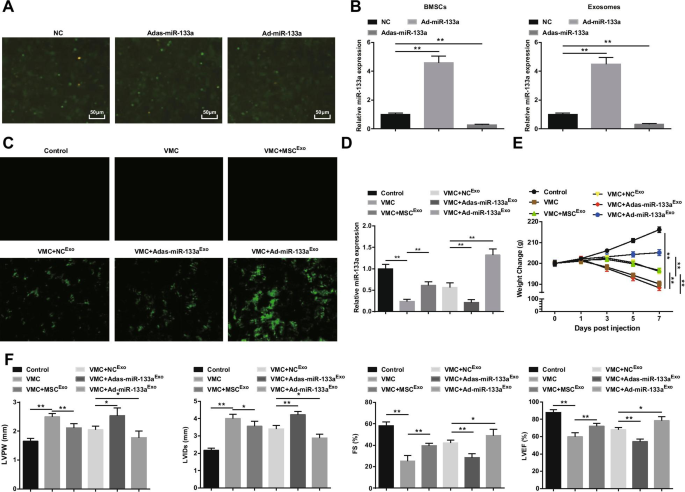
증가된 엑소좀 miR-133a는 심근염의 증상을 개선합니다
miR-133a 재조합 아데노바이러스로 BMSC의 형질감염이 관찰되었다(도 2A). NC, Ad-miR-133a 및 Adas-miR-133a의 많은 녹색 형광 발현이 도립형광현미경에서 관찰되었으며, 이는 재조합 아데노바이러스 벡터가 BMSC를 효과적으로 형질감염시킬 수 있음을 나타낸다. miR-133a의 형질감염 효율을 테스트하기 위해 BMSC 및 엑소좀에서 miR-133a 발현을 RT-qPCR로 측정했습니다. miR-133a 상향 조절은 miR-133a 발현을 증가시키는 반면 miR-133a 하향 조절은 miR-133a 발현을 감소시키는 것으로 밝혀졌습니다(그림 2B). 그 후, 우리는 miR-133a를 포함하는 엑소좀을 쥐에 주입했습니다. 도립형광현미경에서 NC
Exo
처리 후 VMC 쥐에서 녹색 형광 발현이 관찰되었습니다. , Ad-miR-133a
엑소
, 또는 Adas-miR-133a
엑소
, 재조합 아데노바이러스 벡터가 쥐의 심근 조직에 감염되었음을 나타냅니다(그림 2C). RT-qPCR 실험은 또한 VMC 쥐에서 miR-133a 발현이 분명히 감소했음을 발견했습니다. miR-133a 발현은 Ad-miR-133a
Exo
를 주사한 VMC 쥐에서 분명히 증가했습니다. 그러나 Adas-miR-133a
Exo
를 주사한 VMC 쥐에서 감소 (그림 2D). 랫드의 일반적인 상태는 정상 대조군 랫트의 일반적인 상태가 정상이었으며 VMC 랫트와 VMC 랫트에서 Adas-miR-133a
Exo , 거칠고 흐트러진 머리카락, 호흡곤란, 적은 양의 식이요법 등. MSC
Exo
로 처리된 VMC 쥐에서 , NC
엑소
및 Ad-miR-133a
엑소
, 이러한 징후는 다양한 정도로 개선되었습니다. VMC 쥐의 체중은 감염 1일 후부터 지속적으로 감소하였고 MSC
Exo
주입 , NC
엑소
, 또는 Ad-miR-133a
엑소
쥐의 무게를 늘렸습니다. Ad-miR-133a
Exo
로 처리된 VMC 쥐의 체중 Adas-miR-133a
Exo
를 주사한 VMC 쥐의 체중이 분명히 증가했습니다. 분명히 감소했습니다(그림 2E).
<그림>
상향 조절된 엑소좀 miR-133a는 심근염을 완화합니다. A miR-133a 재조합 아데노바이러스의 BMSC 형질감염; 나 miR-133a 조절 후 BMSC 및 엑소좀에서 miR-133a 발현의 RT-qPCR 검출; ㄷ 도립 형광 현미경을 통해 테스트한 miR-133a 형질감염 효율; 디 RT-qPCR을 통해 테스트한 심근 조직에서 miR-133a의 상대적 발현; 이 각 그룹에서 쥐의 체중 변화; F 각 그룹의 쥐에서 LVPW, LVID, FS 및 LVEF의 측정. *피 <0.05; **피 <0.001
심근 기능 관찰은 (그림 2F) VMC 쥐가 LVPW와 LVID를 증가시켰고 FS와 LVEF를 감소시켰음을 시사했습니다. 엑소좀 주입 후, VMC 쥐에서 LVPW 및 LVID가 감소하고 FS 및 LVEF가 분명히 상승했습니다. Adas-miR-133a
엑소
Ad-miR-133a
Exo
동안 치료 장애 VMC 쥐의 심근 기능 향상
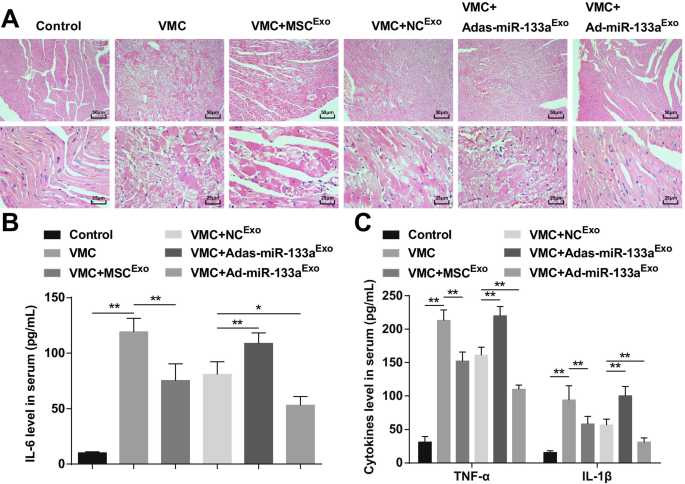
상향 조절된 엑소좀 miR-133a는 VMC 쥐의 심근 조직 염증을 억제합니다.
HE 염색은 정상 대조군 쥐의 심근 섬유가 밀접하게 배열되어 있고 중간엽에 염증 세포 침윤이 없음을 나타냈습니다. VMC 쥐의 심근세포는 와해되었고 중간엽은 다수의 염증 세포에 의해 침윤되었다. MSC
Exo
를 주입한 VMC 쥐의 심근세포 또는 NC
엑소
중간엽에 소량의 염증 세포가 침투하여 질서 정연하게 배열되었습니다. Adas-miR-133a
Exo
후 VMC 쥐의 심근세포 치료가 무질서하게 배열되었고 중간엽의 염증세포가 침윤되었다. Ad-miR-133a
Exo
로 처리된 VMC 쥐의 심근세포 명백한 염증세포 침윤 없이 질서정연하게 배열되었다(Fig. 3A).
<그림>
증가된 엑소좀 miR-133a는 VMC로 심근 조직의 염증을 억제합니다. A 각 그룹의 쥐 심근 조직의 H 염색; 나 ELISA를 통해 테스트된 혈청에서 IL-6의 발현; ㄷ ELISA를 통해 테스트한 혈청 내 TNF-α 및 IL-1β의 발현. *피 <0.05; **피 <0.001
ELISA는 (그림 3B, C) 염증 인자(TNF-α, IL-1β 및 IL-6)가 VMC 쥐에서 분명히 증가했음을 나타냅니다. Ad-miR-133a
Exo
가 주입된 VMC 쥐 염증 인자 수치가 감소했습니다. Adas-miR-133a
엑소
치료는 VMC 쥐에서 염증 인자 상승을 유발했습니다.
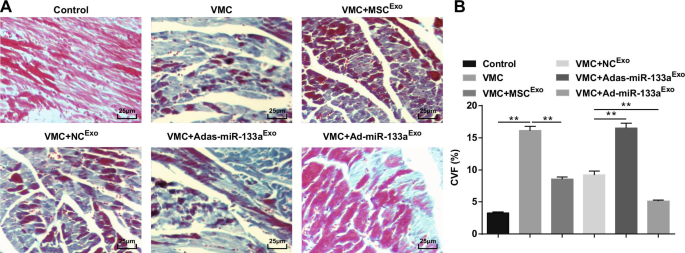
증가된 엑소좀 miR-133a는 VMC가 있는 쥐의 심근 조직에서 CVF를 감소시킵니다.
Masson 염색은 정상 쥐의 심근 섬유가 밀접하게 배열되어 있으며 파란색 콜라겐 섬유가 거의 없음을 보여주었습니다. CVB3 주사 후 심근 세포는 결합 조직 증식과 많은 수의 청색 콜라겐 섬유와 함께 비대했고 CVF는 분명히 상승했습니다. 엑소좀을 처리한 결과 심근세포가 질서정연하게 배열되었고, 세포간 결합조직 증식이 감소하였고, 청색 콜라겐 섬유와 CVF가 뚜렷하게 감소하였다. Adas-miR-133a
Exo
를 주사한 VMC 쥐의 심근 세포간 공간 확장되었고, 세포가 분명히 확대되었으며, 파란색 콜라겐 섬유와 CVF가 분명히 증가했습니다. Ad-miR-133a
Exo
를 투여한 VMC 쥐의 세포간 공간 감소, 청색 콜라겐 섬유 분포 및 CVF 감소 치료(그림 4A, B).
<사진>
상향 조절된 엑소좀 miR-133a는 VMC가 있는 쥐의 심근 조직에서 CVF를 감소시킵니다. A 쥐에서 심근 조직의 Masson 염색; 나 각 그룹에서 쥐의 콜라겐 부피 분율. **피 <0.001
증가된 엑소좀 miR-133a는 VMC가 있는 쥐의 심근 조직에서 콜라겐 I, 콜라겐 III, TGF-β1 및 CTGF의 발현을 감소시킵니다.
콜라겐 I과 콜라겐 III는 콜라겐의 주성분으로 세포 접합부와 세포막, 세포간 물질, 세포질에 주로 분포한다. TGF-β1과 CTGF는 섬유증의 특징적인 단백질입니다. RT-qPCR의 결과는 콜라겐 I, 콜라겐 III, TGF-β1 및 CTGF mRNA 발현 수준이 VMC 쥐에서 증가했지만 엑소좀 처리 후에 감소한다는 것을 보여주었습니다. Ad-miR-133a
Exo
로 처리된 VMC 쥐 Adas-miR-133a
Exo
후 VMC 쥐는 콜라겐 I, 콜라겐 III, TGF-β1 및 CTGF의 mRNA 발현 수준이 감소한 반면 VMC 쥐는 치료는 반대 상황을 보여주었다(그림 5A-C).
Elevated exosomal miR-133a reduces the mRNA expression of collagen I, collagen III, TGF-β1 and CTGF in myocardial tissues of rats with VMC. A Collagen I mRNA expression in myocardial tissues of rats was detected by RT-qPCR; 나 Collagen III mRNA expression in myocardial tissues of rats was detected by RT-qPCR; ㄷ TGF-β1 and CTGF mRNA expression in myocardial tissues of rats was detected by RT-qPCR. *피 < 0.05; **피 < 0.001
Up-regulated Exosomal miR-133a Inhibits the Cardiomyocyte Apoptosis in Myocardial Tissues of Rats with VMC
TUNEL staining showed that the apoptotic cardiomyocytes were brownish black or brownish yellow with nuclear condensation. The number of apoptotic cells was increased in VMC rats which would be attenuated by exosome treatment. The VMC rats injected with Ad-miR-133a
Exo
had reduced number of apoptotic cells and those injected with Adas-miR-133a
Exo
had increased number of apoptotic cells (Fig. 6A, B).
Increased exosomal miR-133a inhibits the cardiomyocyte apoptosis in myocardial tissues of rats with VMC. A TUNEL staining of rat myocardial tissues in each group; 나 The number of TUNEL positive cells in each group. **피 < 0.001
Elevated Exosomal miR-133a Depresses EMT in Myocardial Tissues of Rats with VMC
E-cadherin, α-SMA, and FSP-1 are key indicators of EMT. Results of RT-qPCR demonstrated that α-SMA and FSP-1 mRNA expression levels were elevated and E-cadherin mRNA expression level was decreased in VMC rats. In addition, α-SMA and FSP-1 mRNA expression levels were reduced and E-cadherin mRNA expression level was increased in VMC rats after exosome treatment. α-SMA and FSP-1 mRNA expression levels were elevated and E-cadherin mRNA expression level was decreased in VMC rats treated with Adas-miR-133a
Exo
, while the expression of these indicators was opposite in VMC rats injected with Ad-miR-133a
Exo
(Fig. 7A, B).
Up-regulated exosomal miR-133a represses EMT in myocardial tissues of rats with VMC. A The mRNA expression of α-SMA in rat myocardial tissues in each group was detected by RT-qPCR; 나 The mRNA expression of FSP-1 and E-cadherin in rat myocardial tissues in each group was detected by RT-qPCR. *피 < 0.05; **피 < 0.001
Up-regulated Exosomal miR-133a Depresses Inflammation of Cardiomyocytes in VMC
As a result, fluorescence microscopy captured green fluorescent expression in VMC rats treated with NC
Exo
, Ad-miR-133a
Exo
, or Adas-miR-133a
Exo
, indicating that the recombinant adenovirus vector infected cardiomyocytes of rats (Fig. 8A). RT-qPCR and ELISA discovered that (Fig. 8B, D) miR-133a expression was reduced and inflammatory factors (TNF-α, IL-1β, and IL-6) were increased in VMC rats, which would be reversed by exosome treatment. The VMC rats treated with Ad-miR-133a
Exo
had up-regulated miR-133a and decreased inflammatory factors in VMC rats, while those treated with Adas-miR-133a
Exo
presented decreased miR-133a and increased levels of inflammatory factors in VMC rats.
Elevated exosomal miR-133a restrains inflammation of cardiomyocytes in VMC. A miR-133a transfection efficiency tested via inverted fluorescence microscope; 나 The relative expression of miR-133a in cardiomyocytes of rats in each group; ㄷ IL-6 expression in culture supernatant of cardiomyocytes in each group; D TNF-α and IL-1β expression in culture supernatant of cardiomyocytes in each group. *피 < 0.05; **피 < 0.001
Elevated Exosomal miR-133a Promotes Cell Viability, and Represses Apoptosis of Cardiomyocytes in VMC
The apoptosis and the cell viability were detected via AnnexinV-APC/PI double staining and CCK-8 assay. The results revealed that there was an obvious increase in apoptosis rate, a decrease in cell viability of cardiomyocytes in VMC rats. Exosome treatment reduced apoptosis rate and enhanced the viability of cardiomyocytes. Adas-miR-133a
Exo
enhanced the apoptosis rate and disrupted the viability of cardiomyocytes in VMC rats. Ad-miR-133a
Exo
treatment functioned the opposite effects on cardiomyocytes of VMC rats (Fig. 9A–C).
Increased exosomal miR-133a promotes viability and represses apoptosis in cardiomyocytes in VMC. A The cardiomyocytes apoptosis detected via flow cytometry; 나 Quantification results of A; ㄷ The cell viability detected via CCK-8 assay. *피 < 0.05; **피 < 0.001
miR-133a Targets MAML1
It has been reported that up-regulated miRNA-193b reduces myocardial I/R damage by targeting MAML1 [18]. Based on that, we cross-screened downstream genes of miR-133a through bioinformatics websites PITA, miRanda, PicTar, microT and miRmap, and selected MAML1 as a target of miR-133a (Fig. 10A). We constructed MAML1-wt or MAML1-mut, and co-transfected cardiomyocytes with miR-133a mimic or NC. The results showed that miR-133a mimic reduced the luciferase activity of MAML1-wt (Fig. 10B). The RIP experiment further verified the targeting relationship between miR-133a and MAML1 (Fig. 10C). RT-qPCR and Western blot detection of MAML1 expression showed that MAML1 expression was decreased in cardiomyocytes transfected with miR-133a mimic (Fig. 10D, E).
miR-133a targets MAML1. A miR-133a’s targets predicted on bioinformatics websites; 나 The targeting relationship between miR-133a and MAML1 verified by dual luciferase reporter gene experiment; ㄷ The targeting relationship between miR-133a and MAML1 verified by RIP experiment; D /E MAML1 expression changes after up-regulation of miR-133a detected by RT-qPCR and Western blot. *피 < 0.05; **피 < 0.01; ***피 < 0.001
Inhibition of MAML1 has a Protective Effect on Rats with Myocarditis and Reverses the Effect of miR-133a-Inhibited Exosomes on Rats with VMC
To further study the effect of miR-133a-regulated MAML1 on rats with VMC, we injected si-MAML1 or si-NC adenovirus into VMC rats or VMC rats that had been treated with miR-133a-silenced exosomes. The injection success was validated by RT-qPCR and Western blot (Fig. 11A, B). The results manifested that injection of si-MAML1, the weight of VMC rats was increased (Fig. 11C), cardiac function was improved (Fig. 11D–G), myocardial tissue pathology and fibrosis were attenuated (Fig. 12A–C), serum inflammation (Fig. 12D, E) and cardiomyocyte apoptosis (Fig. 13A–G) were inhibited. Also, the deleterious effects of miR-133a-silenced exosomes in VMC rats were reversed after injection of si-MAML1.
Inhibition of MAML1 has a protective effect on myocarditis rats and can reverse the effect of miR-133a-silenced exosomes on rats with VMC. A /B MAML1 expression in myocardial tissue of rats detected by RT-qPCR and Western blot; C. Weight change of rats; D –G Determination of LVPW, LVIDs, FS and LVEF in rats; **피 < 0.01; ***피 <0.001; ****P < 0.0001
Inhibition of MAML1 can reverse the effect of miR-133a-silenced exosomes on rats with VMC. A HE staining of rat myocardial tissue; 나 Masson staining of myocardial tissues in rats; ㄷ CVF of rats; D The expression of IL-6 in serum tested via ELISA; E The expression of TNF-α and IL-1β in serum tested via ELISA.****P < 0.0001
Inhibition of MAML1 can reverse the effect of miR-133a-inhibiting exosomes on rats with VMC. A Collagen I mRNA expression in myocardial tissues of rats was detected by RT-qPCR; 나 Collagen III mRNA expression in myocardial tissues of rats was detected by RT-qPCR; ㄷ TGF-β1 and CTGF mRNA expression in myocardial tissues of rats was detected by RT-qPCR; D TUNEL staining of rat myocardial tissues in each group; E The number of TUNEL positive cells in each group. F The mRNA expression of α-SMA in rat myocardial tissues in each group was detected by RT-qPCR; G The mRNA expression of FSP-1 and E-cadherin in rat myocardial tissues in each group was detected by RT-qPCR. *피 < 0.05; **피 < 0.01; ****P < 0.0001
Discussion
Myocarditis is an inflammatory heart illness resulting in DCM and heart failure and is most frequently induced by viral infections such as CVB3 [2]. A study has revealed that miR-133 relieves cardiomyocyte apoptosis and electrical remodeling in mice with VMC [23]. Additionally, changed exosomal miRNAs are also found to be linked with the pathogenesis of CVB3-induced myocarditis [14]. Exosomes derived from cardiac progenitor cells ease CVB3-induced apoptosis via restraining the proliferation of CVB3 in VMC [24]. This study explored the regulatory mechanism of BMSC-derived exosomal miR-133 on myocardial fibrosis and EMT in VMC rats (Additional file 1:Fig. 1).
The study found that the expression of miR-133a was decreased in VMC. As demonstrated before, miR-133a expression is decreased in MI [7]. A study has also suggested that the relative expression of miR-133 in mouse hearts of the VMC is obviously decreased with contrast to the controls [23]. There are some connections of miRNAs with exosomes. The differential expression of exosomes and of exosomal miRNAs in illness has been regarded as biomarkers of disease with performance of noninvasive clinical diagnosis together with their therapeutic potentials [25]. Lin et al. have found that miR-133 is specially sorted into hypoxia/reoxygenation (H/R)-caused human endothelial progenitor cells-derived exosomes to increase fibroblast angiogenesis and EMT [26]. Another study has revealed that MSCs exhibits a communication with brain parenchymal cells and may modulate neurite outgrowth by transfer of miR-133b to neural cells via exosomes [27].
The major finding of this work manifested that up-regulated exosomal miR-133a promoted cell viability, inhibited inflammation, apoptosis, EMT, and fibrosis in rats with VMC. They suits well with a former research that miR-133a silence reverses the Astragalus polysaccharides treatment-induced osteosarcoma MG63 cell proliferation inhibition, together with cell apoptosis promotion [28]. Another study has revealed that overexpressed miR-133a suppresses angiogenesis, apoptosis, fibrosis, and inflammation, while accelerating therapeutic cardiac remodeling in ischemic myocardial illnesses [29]. Similar to our study, Li et al. have stated that miR-133 inhibits cardiomyocyte apoptosis by regulating the expression of apoptosis-related genes in the hearts of VMC mice [23]. The over-expressed miR-133a has been reported to depress hypoxia-induced apoptosis and strengthen cardiomyocyte survival [30]. Meanwhile, the up-regulated serum exosomal miR-30a and miR-181d may have the potentials to be applied as biomarkers for VMC diagnosis [14].
Another finding in our study was that up-regulated exosomal miR-133a decreased CVF, reduced the expression of collagen I and collagen III in rats with VMC. A article has elucidated that released fibroblast growth factor-18 from a collagen membrane causes osteoblastic activity participating in down-regulated miR-133a [31]. In vitro excessive expression of miR-133a depresses cardiomyocyte hypertrophy and reduces collagen expression [32], as evidenced in another study. CVF equals the ratio of collagen area to the sum of myocardial area and collagen area, and the mean value shows the CVF of the section [33]. This finding is also reported by Wang et al. that VMC mice model is successfully constructed by CVB3 infection, manifesting apparent higher CVF expression in contrast with the control group [34]. Moreover, the finding is consistent with that of Ferreira et al. who demonstrates that miR-133a may take on a major role in the modulation of gene expression in chronic Chagas disease cardiomyopathy pathogenesis, with potential link as diagnostic and prognostic tools [8]. Furthermore, evidence has shown that knocking down MAML1 can reduce the hypertrophy of pre-treated cardiomyocytes [35]. In our study, we found that MAML1 was the target gene of miR-133a and inhibition of MAML1 reversed the effects of miR-133a-silenced exosomes on rats with VMC. In myocardial ischemia–reperfusion injury, miR-193b-mediated down-regulation of MAML1 could in part reduce infarction and myocardial enzymes, as well as attenuate apoptosis of cardiomyocytes [18]. Also, there is a report suggesting that deficiency of MAML1 could relieve hepatic fibrogenesis [19].
Conclusion
In conclusion, this present study offers evidence that miR-133a is down-regulated in rats with VMC, and elevated exosomal miR-133a improves cardiac function and restrains myocardial fibrosis and EMT in rats with VMC, as well as enhances viability and represses apoptosis of cardiomyocytes in VMC through targeting MAML1. Our study also suggests that inhibition of MAML1 has a protective effect on rats with myocarditis and reverses the effect of miR-133a-inhibited exosomes on rats with VMC. The identification of the exosomal miR-133a in myocardial fibrosis and EMT of myocarditis may potentially widen our understanding of mechanisms underpinning myocarditis and also bear clinical value as a novel molecular target. More researches should be undertaken for making inroads into the treatment of this disease.
약어
miR-133:
MicroRNA-133
BMSC-Exo:
Bone marrow mesenchymal stem cell-derived exosome
EMT:
Epithelial–mesenchymal transition
VMC:
Viral myocarditis
CVF:
Collagen volume fraction
DCM:
Dilated cardiomyopathy
CVB3:
Coxsackie B3 virus
miRNAs:
MicroRNAs
MI:
Myocardial infarction
SPF:
Specific pathogen-free
SD:
Sprague–Dawley
DMEM:
Dulbecco’s Modified Eagle Medium
PBS:
인산염 완충 식염수
FBS:
태아 소 혈청
NC:
Negative control
MOI:
Multiplicity of infection
LVPW:
Left ventricular posterior wall thickness
LVIDs:
Left ventricular end-systolic diameter
LVEF:
Left ventricular ejection fraction
HE:
헤마톡실린-에오신
TUNEL:
Terminal deoxynucleotidyl transferase-mediated deoxyuridine triphosphate-biotin nick end-labelin