복구된 microRNA-326-5p는 뇌허혈/재관류 손상에서 STAT3 억제를 통해 신경 세포 사멸을 억제하고 미토콘드리아 손상을 약화시킵니다.
초록
연구는 대뇌 허혈/재관류 손상(CI/RI)에서 microRNA(miRNA)의 역할을 크게 탐구했습니다. 그러나 CI/RI에서 miR-326-5p의 특정 메커니즘은 여전히 파악하기 어렵습니다. 따라서, 이 연구는 CI/RI에서 miR-326-5p/신호 변환기 및 전사-3(STAT3) 축의 활성제의 메커니즘을 밝히는 것이었습니다. 두 가지 모델(일차 쥐 피질 뉴런의 산소 및 포도당 결핍[OGD] 및 Sprague-Dawley 쥐의 중간 대뇌 동맥 폐색[MCAO])은 각각 시험관 내 및 생체 내에서 CI/RI를 모방하기 위해 설정되었습니다. 기능 상실 및 증가 분석은 OGD 처리된 뉴런과 MCAO 쥐로 수행되었습니다. 그 후, OGD 처리된 뉴런의 생존력, 세포 사멸, 산화 스트레스 및 미토콘드리아 막 전위뿐만 아니라 MCAO 쥐의 뇌 조직에서 병리학적 변화, 세포 사멸 및 미토콘드리아 막 전위를 테스트했습니다. Mitofusin-2(Mfn2), miR-326-5p 및 STAT3 발현이 OGD 처리된 뉴런과 MCAO 쥐의 뇌 조직에서 검출되었습니다. Mfn2 및 miR-326-5p는 감소되었고 STAT3는 MCAO 쥐의 OGD 처리된 뉴런과 뇌 조직에서 증가했습니다. miR-326-5p는 STAT3 발현을 표적화하고 부정적으로 조절합니다. miR-326-5p를 복원하거나 STAT3를 감소시키면 생존력이 강화되고 세포자멸사 및 산화 스트레스가 억제되며 미토콘드리아 막 전위가 증가하고 OGD 처리된 뉴런에서 Mfn2 발현이 증가합니다. miR-326-5p를 상향 조절하거나 STAT3를 하향 조절하면 MCAO가 있는 쥐의 뇌 조직에서 병리학적 변화를 완화하고 세포자멸사를 억제하고 미토콘드리아 막 전위와 Mfn2 발현을 증가시켰습니다. 이 연구는 상향 조절된 miR-326-5p 또는 하향 조절된 STAT3가 Mfn2 발현을 증가시켜 CI/RI를 보호한다는 것을 설명합니다.
소개
뇌허혈/재관류 손상(CI/RI)은 일종의 뇌 손상으로 허혈성 뇌졸중이 뒤따른다[1]. 대뇌 I/R의 가장 특징적인 특징은 재관류 후 초기 일과성 대뇌 허혈이다[2]. CI/RI는 신경 세포 사멸을 활성화하고 해마 및 피질 손상을 유발합니다[3]. 현재 가장 일반적인 임상 적용은 CI/RI에 대한 혈전 용해제입니다[4]. 그러나 재관류는 반응성 산소종(ROS)의 생성을 증가시켜 세포 내 DNA 손상, 산화 스트레스 관련 손상, 단백질 산화 및 지질 과산화를 유발하여 혈액-뇌 장벽 및 부종을 더욱 악화시킬 것입니다[5]. 따라서 CI/RI의 우선 순위는 새로운 대상 옵션을 검색해야 하는 시급함입니다.
MicroRNA(miRNA)는 CI/RI에서 잠재적 및 예후 역할로 광범위하게 고려됩니다[6, 7]. 예를 들어, miR-202-5p가 허혈성 손상에 있는 중대뇌동맥폐쇄(MCAO) 모델 쥐의 산소 및 포도당 결핍(OGD) 유발 세포 손상뿐만 아니라 신경 손상 및 신경학적 결손을 약화시킨다는 기사가 있습니다[8] . 또한 miR-98의 상향 조절은 I/R 뇌졸중[9]이 있는 마우스에서 신경학적 결과를 개선하고 miR-451의 과발현은 CI/RI[10]가 있는 마우스에서 허혈성 대뇌 세포자멸사를 완화하는 것으로 밝혀졌습니다. 구체적으로, miR-326-5p는 내피 전구 세포에 대한 혈관신생 촉진 능력을 가지며 급성 심근경색 후 심장 기능을 향상시킬 수 있습니다[11]. 그러나 CI/RI에서 miR-326-5p를 중심으로 하는 연구는 아직 초기 단계입니다. 신호 변환기와 전사 활성제(STAT)는 CI/RI에 의해 활성화되는 독특한 단백질 군입니다[12]. STAT3는 생물정보학 웹사이트에서 miR-326-5p의 표적으로 예측되었습니다. 따라서 우리는 CI/RI에서 miR-326-5p 매개 STAT3의 역할을 연구했습니다. STAT3는 염증 유발 매개체를 방출하여 허혈성 신경 염증 과정과 이차적 뇌 손상을 악화시킵니다[13, 14]. 구체적으로, Janus-activated kinase 2(JAK2)/STAT3 경로는 CI/RI에 의해 유도된 세포 사멸을 차단하는 기능적 역할을 한다[15]. Mitofusin-2(Mfn2)는 미토콘드리아 융합 인자로 심장-뇌혈관 I/RI[16]를 보호하고 허혈성 뇌 손상에서 저산소증으로 인한 신경 세포 사멸을 개선할 수 있습니다[17]. Mfn2가 CI/RI에 보호 효과를 발휘하는 것으로 기록되어 있습니다[18].
한마디로, 덜 심층적인 조사는 CI/RI에서 miR-326-5p와 STAT3의 결합된 역할을 발견했습니다. 이를 감안할 때 이 연구는 miR-326-5p가 STAT3를 표적으로 하여 CI/RI를 감쇠한다는 가설로 시작되었습니다.
자료 및 방법
윤리 성명서
실험은 Yangzhou 대학 부속 병원의 동물 관리 및 사용 위원회의 승인을 받았습니다. Yangzhou University, 그리고 그것은 Yangzhou University의 부속 병원의 실험용 쥐의 관리 및 사용에 대한 가이드와 함께 수행되었습니다. 양주대학교.
실험 동물
Yangzhou University (Yangzhou, China)의 비교 의학 센터의 수컷 성체 Sprague-Dawley 쥐(6-8주령, 250 ± 30g)는 음식과 물에 자유롭게 접근할 수 있는 특정 병원체 없는 환경에서 유지되었습니다. (24 ± 1)°C, (50 ± 5)% 습도 및 12시간 명암 주기
세포 분리 및 식별
쥐를 참수로 안락사시키고 뇌를 1mm
3
로 절단했습니다. 0.4% 트립판 블루 용액과 9:1(트립판 블루의 최종 농도는 0.04%) 혼합하여 세포 현탁액을 구성하고 혈구계로 세포 밀도 및 생존율을 계산했습니다. 셀(1 × 10
6
세포/mL)를 5 × 10
6
에서 L-폴리리신(0.1 mg/mL)으로 사전 코팅된 세포 배양 플라스크에 접종했습니다. 플라스크당 세포. 세포가 벽에 부착되면 재생된 배지(NeurobasalA + B27 + L-glutamine)와 함께 배양하고 2-3일마다 배지를 반으로 교체하면서 6-7일 동안 계속 배양했습니다.
세포 식별:랫트 피질 뉴런을 사용하여 1차 토끼 항-쥐 네스틴(NES) 폴리클로날 항체(1:200) 및 플루오레세인 이소티오시아네이트(FITC) 표지된 염소 항-토끼 면역글로불린과 함께 인큐베이션된 뉴런 슬라이드를 준비했습니다. G(1:50, 둘 다 Cell Signaling Technology, Beverly, MA, USA). 그 후 세포를 형광소광제로 봉인하고 형광현미경으로 관찰하였다.
OGD 모델 수립
뉴런은 포도당이 없는 Earle's 용액에서 배양되었고 95% N2의 혼합 가스에 노출되었습니다. 및 5% CO2 . 30분의 양압 환기 후, 세포가 배양되는 저산소 환경이 형성되었습니다(시험관 내 OGD에 의한 허혈 시뮬레이션). 배양 90분 후, 무글루코스 Earle's 용액을 제거한 후 유지 세포 배지와 함께 세포를 배양하였다. 세포는 후속 실험(시험관 내 산소-포도당 재관류에 의한 재관류 시뮬레이션)을 위해 일상적으로 배양되었습니다. 정상 산소 상태에서 정상 배지가 대조군으로 사용되었습니다.
뉴런을 대조군으로 나누었습니다. OGD 그룹, 음성 대조군(NC) 그룹(스크램블된 올리고뉴클레오티드로 형질감염된 OGD 처리 뉴런), miR-326-5p agomir 그룹(miR-326-5p agomir로 형질감염된 OGD 처리 뉴런), sh-STAT3 그룹 (STAT3 shRNA로 형질감염된 OGD 처리 뉴런) 및 miR-326-5p agomir + 과발현(oe)-STAT3 그룹(miR-326-5p agomir 및 pcDNA-STAT3 벡터로 형질감염된 OGD 처리 뉴런). 올리고뉴클레오티드 및 벡터는 모두 GenePharma(중국 상하이)에서 제공했습니다.
리포펙타민 2000(Invitrogen; Thermo Fisher Scientific)을 사용하여 스크램블된 올리고뉴클레오티드, miR-326-5p agomir, STAT3 shRNA 또는 miR-326-5p agomir 및 pcDNA-STAT3를 일차 쥐 뉴런에 형질감염시키고 역전사 정량에 의해 형질감염 효능을 시험했습니다. 48시간 후 중합효소 연쇄 반응(RT-qPCR) 또는 웨스턴 블롯.
3-(4,5-디메틸티아졸-2-일)-2,5-디페닐테트라졸륨 브로마이드(MTT) 분석
뉴런은 1 × 10
6
에서 정지되었습니다. 세포/mL 및 각각 0, 24 및 48시간 동안 배양되었습니다. 멸균 환경에서 뉴런을 10% MTT 용액에서 2-4시간 동안 배양하고 결정이 충분히 용해될 때까지 150μL Formanzan 용해 용액(Dimethyl Sulfoxide)을 첨가했습니다. 광학 밀도(OD) 값은 570 nm에서 마이크로플레이트 리더에서 측정되었습니다.
유세포분석 세포자멸사 분석
뉴런을 0.25% 트립신으로 분리하고 2000rpm에서 원심분리하고 PBS에 재현탁했습니다. 그런 다음, 뉴런을 2000 rpm에서 원심분리하고, 펠렛을 결합 완충액에 의해 재현탁하고 5 μL Annexin V-FITC 및 5 μL propidium iodide(PI)와 함께 연속적으로 인큐베이션했습니다. 세포자멸사 비율을 감지하기 위해 유세포 분석기를 적용했습니다.
JC-1 미토콘드리아 막 전위 검출
6웰 플레이트의 세포를 PBS로 헹구고 1mL 세포 배양 배지 및 1mL JC-1 염색 작업 용액(50μL JC-1, 8mL 초순수 및 2mL JC-1 염색 완충액)을 첨가했습니다(SolarbioScience , Beijing, China) 및 15분 동안 인큐베이션했습니다. 1 × JC-1 염색 완충액은 증류수 4mL와 JC-1 염색 완충액 1mL로 구성하였다. 인큐베이션 후, 세포를 트립신 처리하고 JC-1 완충액에 재현탁하고 유세포 분석기에서 테스트했습니다. 녹색 형광의 상대적 비율을 계산했습니다.
산화 스트레스 감지
세포 내 ROS 수준은 2,7-디클로로플루오레세인 디아세테이트(DCF-DA) 검출 키트(Abcam)로 측정했습니다. 요컨대, 뉴런을 0.25% 트립신으로 분리하고 재현탁하고 30분 동안 10μm DCF-DA와 함께 인큐베이션했습니다. 그 후 495 nm/529 nm에서 여기/방출광을 이용한 DCF 형광을 형광 분광법(BD Biosciences)으로 측정했습니다.
세포내 malondialchehyche(MDA), 글루타티온(GSH) 함량 및 슈퍼옥사이드 디스뮤타제(SOD) 활성의 검출은 상업용 분석 키트(Beyotime Biotechnology Co., Shanghai, China)를 사용하여 화학적 비색법에 의해 검출되었습니다. OD 값은 마이크로플레이트 리더로 측정했습니다.
쥐 모델 확립
펜토바르비탈 나트륨(50㎍/kg)을 복강내 주사하여 쥐를 마취시켰다. 목에 정중선 절개를 하고 외부 경동맥을 결찰하고 총경동맥을 동맥 클립으로 막았다. 3-0 실리콘 코팅된 모노필라멘트 나일론 봉합사를 내경동맥에 내경동맥에 내광성이 있을 때까지 조심스럽게 삽입했습니다. 일과성 뇌허혈 90분 후 봉합사를 제거하고 다시 24시간 동안 재관류를 시행하였다. 가짜 그룹은 봉합사가 내부 경동맥에 들어가지 않는다는 점을 제외하고는 동일한 처리를 받았습니다. 수술 중 직장 온도를 37 ± 0.5 °C로 유지하기 위해 항온 침대를 사용했습니다[19].
래트를 6개 그룹(n =6)으로 할당하고 MCAO 모델을 수립하기 전에 플라스미드 또는 miR을 측면 뇌실에 주입했습니다. 5 μL 스크램블된 올리고뉴클레오티드 [100 μM]), miR-326-5p agomir 그룹(5 μL miR-326-5p agomir[100 μM]의 뇌실 측면 주사를 가진 모델링된 쥐), sh-STAT3 그룹(모델링된 쥐 5 μL STAT3 간섭 벡터[100 μM]) 및 miR-326-5p agomir + oe-STAT3 그룹의 측면 뇌실 주사 STAT3 과발현 벡터 [100 μM]) [20, 21].
표본 수집
쥐는 마취에 의해 안락사되었고, 심장은 관류되었고, 전체 뇌가 얻어졌다. 뇌 손상 조직을 1 × 1 × 1 mm
3
로 절단했습니다. 파라핀 절편 준비, RT-qPCR, 웨스턴 블롯 분석 및 미토콘드리아 막 전위 검출을 위한 조직 덩어리
대뇌 피질의 미토콘드리아 막 전위 감지
SD 쥐의 대뇌 피질을 분리하여 1mm
3
로 절단했습니다. . 조직 블록을 300 메쉬 나일론 망에 놓고 PBS를 첨가했습니다. 그 다음, 세포 현탁액을 1000 r/min으로 원심분리하고, 70% 에탄올을 첨가하고, 다시 1000 r/min으로 원심분리하였다. 그 후, 샘플을 PBS 1mL에 재현탁하고 현탁액(1 × 10
6
세포, 100μL)을 Rhodamine123 염료 용액(10μL, 5μg/mL)과 반응시키고 488nm에서 유세포 분석기로 분석했습니다.
HE 염색
쥐의 뇌 조직을 4% 파라포름알데히드에 24시간 동안 담그고 파라핀 절편을 준비하고 4μm로 자르고 40°C 물에 펴서 60°C에서 굽습니다. 정기적으로 탈랍 및 수화한 파라핀 섹션을 HE 염색 용액으로 염색했습니다. HE 염색 후 손상된 뉴런은 핵의 수축, 세포 부종, 공포화 및 어두운 핵을 나타냈다. 광학현미경(Nikon, Japan)을 사용하여 이미지와 생존 뉴런 수(mm
2
) 허혈성 피질의 수를 세었습니다.
튜넬 염색
파라핀 섹션은 100%, 95%, 80% 및 70% 알코올로 탈랍 및 탈수되었습니다. 그런 다음 절편을 4% 파라포름알데히드에 담그고 0.1% Triton X-100 구연산 나트륨 완충액과 20분 동안, TUNEL 반응 혼합물과 1-1.5시간 동안 배양했습니다. 다음으로, 섹션은 퍼옥시다제 용액 및 디아미노벤지딘(DAB)으로 전개되고, 헤마톡실린으로 대조염색되고, 구배 알코올로 탈수되고, 투과되고, 중성 검으로 밀봉됩니다. TUNEL 양성 세포를 계수하기 위해 광학 현미경으로 절편을 관찰하였다. 핵의 갈색을 띤 노란색 입자는 TUNEL 양성 세포(아폽토시스 세포)로 정의되었습니다.
면역조직화학
섹션은 탈랍, 수화 및 시트레이트 항원에 의해 회수되었으며 내인성 카탈라아제는 3% H2에 의해 차단되었습니다. O2 . 각 절편에 염소 혈청(50μL)을 첨가하고, 1차 항체(50μL)와 함께 배양하고, 2차 항체(50μL) 및 양고추냉이-과산화효소 표지된 스트렙타비딘(50μL)과 반응시켰다. 섹션은 DAB에 의해 전개되었고 헤마톡실린에 의해 대조염색되었으며, 구배 알코올 탈수, 자일렌 투과 및 중성 검 밀봉이 뒤따랐습니다. 절편은 현미경으로 관찰하였다(허혈성 뇌 조직의 세포질 또는 핵은 황색 또는 황갈색 입자였다). OD 값을 측정하고 평균했습니다.
RT-qPCR
조직 및 세포에서 총 RNA 추출은 Trizol(Invitrogen)에 의해 수행되었습니다. mRNA 발현은 SYBR Green PCR Mix(Applied Biosystems, CA, USA)와 QuantStudio TM 6Flex real-time PCR system(Applied Biosystems)을 사용하여 qPCR을 통해 분석하였다. 특정 줄기 루프 역전사 프라이머 및 정방향/역방향 프라이머(BioTNT, Shanghai, China)는 miRNA 수준을 분석하기 위해 설계되었습니다. 표 1은 모든 프라이머를 나열했습니다. 2
−△△CT
방법은 miRNA 또는 mRNA 수준을 계산하기 위해 채택되었습니다.
서부 얼룩 분석
단백질 샘플을 10% 소듐 도데실 설페이트 폴리아크릴아미드 겔 전기영동에 의해 분리하고 폴리비닐리덴 플루오라이드(PVDF) 막에 전기블롯팅하였다. 그 후, PVDF 막을 Tris 함유 식염수에 5% 탈지분유로 차단하고, 1차 항체로 프로브하고, 이차 항체로 2시간 동안 다시 프로브하였다. 그런 다음, 단백질 밴드를 Nitroblue tetrazolium chloride와 5-Bromo-4-Chloro-3-Indolyl Phosphate를 포함하는 알칼리성 인산 가수분해 효소 완충액으로 분석하고 ImageJ 소프트웨어(NIH, Bethesda, MD, USA)로 정량화했습니다. 주요 항체는 STAT3(1:1000), Mfn2(1:1000, Abcam, MA, USA) 및 glyceraldehyde-3-phosphate dehydrogenase(GAPDH)(1:1000, Cell Signaling Technology, Beverly, MA, USA)였습니다.
이중 루시퍼라제 리포터 유전자 분석
miR-326-5p 야생형(WT) 또는 돌연변이체(Mut) 결합 부위를 포함하는 STAT3 3'UTR을 STAT3-WT 및 STAT3-Mut로 명명된 pmirGLO 벡터(Promega, WI, USA)에 클로닝했습니다. 세포를 2 × 10
4
로 24웰 플레이트에 접종했습니다. 세포/웰 및 100ng의 STAT3-WT 또는 STAT3-Mut 루시퍼라제 벡터 및 50nM miR-326-5p agomir 또는 it agomir NC로 70% 컨플루언스로 공동 형질감염되었습니다. Luciferase 활성은 Dual-Luciferase Reporter Assay System(Promega Corporation, WI, USA)을 사용하여 측정되었습니다.
통계 분석
데이터 분석에는 SPSS 21.0(IBM, NY, USA) 통계 소프트웨어를 사용하였다. 데이터는 평균 ± 표준편차(SD)로 표현하였다. 두 그룹 간의 차이는 Student's t-test로 분석한 반면 다중 비교는 일원 분산 분석(ANOVA)으로 분석한 후 Tukey의 사후 테스트로 분석했습니다. 피 양측 검정을 나타내고 P <0.05는 통계적으로 유의미한 것으로 간주되었습니다.
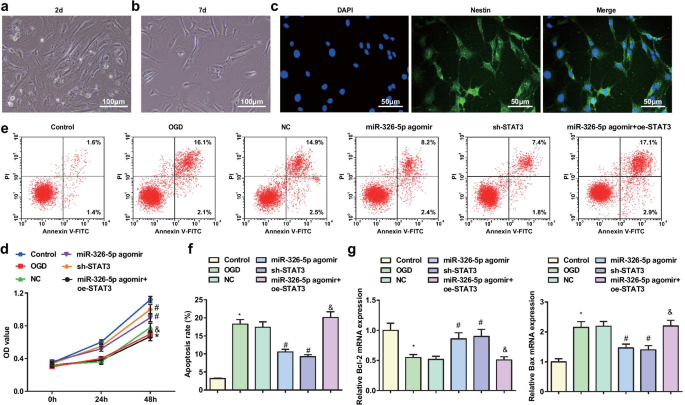
일주일 동안 배양한 쥐의 피질 뉴런은 분화되고 성숙되었습니다. 현미경으로 볼 때 뉴런은 더 큰 뉴런체, 더 투명한 세포질, 밀도가 낮은 큰 핵, 강한 굴절률을 보였다. 뉴런 몸체는 원추형 또는 원형이었고 수상돌기는 엇갈려 벽에 부착되었습니다(그림 1a, b). NES는 면역형광 분석을 통해 높은 특이성을 갖는 뉴런을 찾기 위한 대뇌 뉴런의 특이적 화학적 마커였다. 뉴런은 NES에 특이적으로 결합하는 것으로 밝혀졌으며 녹색 형광을 보였다(그림 1c).
<그림>
miR-326-5p의 상향 조절 또는 STAT3의 하향 조절은 OGD 처리된 피질 뉴런의 세포자멸사를 약화시킵니다. 아 2-d 부착 배양이 있는 대뇌 뉴런(스케일 막대:100μm); ㄴ 7일 부착 배양이 있는 대뇌 뉴런(스케일 막대:100μm); ㄷ NES 면역형광 분석에 의해 염색된 7일 부착 배양을 갖는 대뇌 뉴런(스케일 막대:50 μm); d MTT 분석에 의한 OGD 처리된 피질 뉴런의 생존력; 이 유세포 분석에 의해 검출된 OGD 처리된 피질 뉴런의 세포자멸사; 에 OGD 처리된 피질 뉴런의 세포자멸사 비율; 지 OGD 처리된 피질 뉴런에서 Bcl-2 및 Bax mRNA 발현; *피 대조군과 비교하여 <0.05; #피 NC 그룹과 비교하여 <0.05; &피 <0.05 miR-326-5p agomir 그룹과 비교
MTT 분석은 피질 뉴런 생존력을 감지하기 위해 적용되었습니다(그림 1d). 결과는 쥐 피질 뉴런 생존력이 대조군과 miR-326-5p agomir 그룹에 비해 OGD 그룹과 miR-326-5p agomir + oe-STAT3 그룹에서 손상되었지만 miR-326-5p에서는 향상되었음을 보여주었습니다. agomir 그룹 및 sh-STAT3 그룹 대 NC 그룹(모두 P <0.05).
쥐 피질 뉴런 세포 사멸은 유세포 분석에 의해 결정된 반면 Bcl-2 및 Bax mRNA 발현은 RT-qPCR에 의해 결정되었습니다(그림 1e-g). 그 결과, 대조군 및 miR-326-5p agomir 그룹과 비교하여 OGD 그룹과 miR-326-5p agomir + oe-STAT3 그룹은 세포 사멸률 증가, Bcl-2 발현 감소, Bax 발현 증가를 보였다. (모든 P <0.05). 대조적으로 NC 그룹에서는 miR-326-5p agomir 그룹과 sh-STAT3 그룹에서 세포자멸사 비율이 낮아지고 Bcl-2 수치가 상승했으며 Bax 수치가 억제되었습니다(모든 P <0.05).
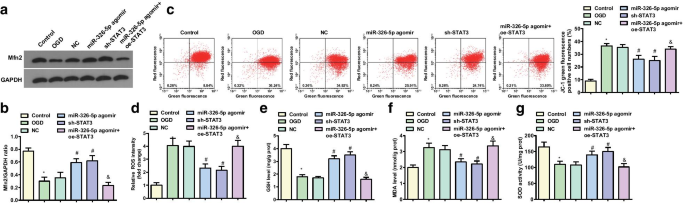
miR-326-5p의 상향 조절 또는 STAT3의 하향 조절은 OGD 손상 후 피질 뉴런에서 Mfn2 및 미토콘드리아 막 전위 수준을 증가시킵니다.
OGD 처리된 피질 뉴런에서의 Mfn2 발현은 웨스턴 블롯 분석에 의해 검출되었다(도 2a, b). Mfn2 단백질 발현은 대조군과 miR-326-5p agomir 군에서 OGD 군과 miR-326-5p agomir + oe-STAT3 군에서 감소한 반면 miR-326-5p agomir 군과 sh에서는 증가하였다. -STAT3 그룹 대 NC 그룹(모두 P <0.05).
<그림>
miR-326-5p의 상향 조절 또는 STAT3의 하향 조절은 OGD 처리된 피질 뉴런에서 Mfn2 및 미토콘드리아 막 전위 수준을 증가시킵니다. 아 , b OGD 처리된 피질 뉴런에서의 Mfn2 단백질 발현; ㄷ OGD 처리된 피질 뉴런에서 미토콘드리아 막 전위 수준의 JC-1 유세포 분석; d miR-326-5p를 상향 조절하거나 STAT3를 하향 조절한 후 OGD 처리된 피질 뉴런의 ROS 함량; 이 miR-326-5p를 상향 조절하거나 STAT3를 하향 조절한 후 OGD 처리된 피질 뉴런에서의 GSH 활성; 에 miR-326-5p를 상향 조절하거나 STAT3를 하향 조절한 후 OGD 처리된 피질 뉴런의 MDA 함량; 지 miR-326-5p를 상향 조절하거나 STAT3를 하향 조절한 후 뉴런의 SOD 활성; *피 대조군과 비교하여 <0.05; #피 NC 그룹과 비교하여 <0.05; &피 <0.05 miR-326-5p agomir 그룹과 비교
JC-1에 의해 검출된 미토콘드리아 막 전위는 대조군 및 miR-326-5p 아고미르 그룹에 대하여 미토콘드리아 막 전위 수준이 OGD 그룹 및 miR-326-5p 아고미르에서 감소함을 발견하였다(도 2c, d) + oe-STAT3 그룹(모든 P <0.05). NC 그룹에 비해 miR-326-5p agomir 그룹과 sh-STAT3 그룹에서 미토콘드리아 막 전위 수준이 증가했습니다(모두 P <0.05).
OGD 처리된 대뇌피질 뉴런에서 ROS 및 MDA 함량, GSH 및 SOD 활성을 측정한 결과 대조군과 miR-326-5p agomir 그룹에 비해 OGD 그룹과 miR-326-5p agomir + oe -STAT3 그룹은 증가된 ROS 및 MDA 함량과 손상된 GSH 및 SOD 활성을 나타냈습니다(모든 P <0.05). NC 그룹과 비교하여 miR-326-5p agomir 그룹과 sh-STAT3 그룹은 ROS 및 MDA 함량이 감소하고 GSH 및 SOD 활성이 강화된 것으로 나타났습니다(모든 P <0.05) (그림 2e–g).
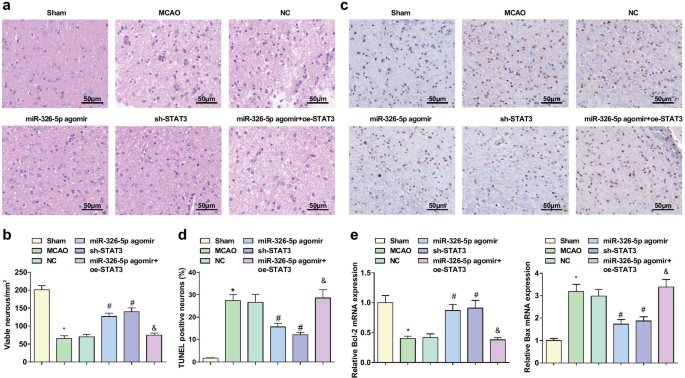
miR-326-5p의 상향 조절 또는 STAT3의 하향 조절은 피질 뉴런의 병리학적 손상을 방해하고 CI/RI가 있는 쥐의 세포자멸사를 억제합니다.
뇌 조직의 병리학적 손상을 관찰하기 위해 H 염색법을 채택한 결과(Fig. 3a, b), sham 그룹에서 뉴런은 구조가 정상이고 옅은 빨간색 세포질, 파란색 핵 및 맑은 핵소체로 깔끔하게 배열되어 있음을 보여줍니다. MCAO군, NC군, miR-326-5p agomir + oe-STAT3군에서는 괴사를 제외하고는 신경세포가 무질서하게 배열되어 더 어둡게 염색되어 핵막 파열, 세포 구조 소실, 핵융합, 깊은 다량의 염색된 핵 및 용해. miR-326-5p agomir 군과 sh-STAT3 군에서 신경세포 종기가 완화되고 신경세포가 질서정연하게 배열되었으며 괴사세포가 감소하여 MCAO군, NC군에 비해 양호한 상태를 나타냈다. <그림>
miR-326-5p의 상향 조절 또는 STAT3의 하향 조절은 피질 뉴런의 병리학적 손상을 방해하고 CI/RI가 있는 쥐의 세포 사멸을 억제합니다. 아 HE 염색에 의해 관찰된 MCAO 래트에서 뇌 조직의 병리학적 손상(스케일 바:50μm); ㄴ miR-326-5p를 상향 조절하거나 STAT3를 하향 조절한 후 MCAO 쥐 뇌 조직의 온전한 뉴런의 수; ㄷ TUNEL 염색에 의해 검출된 MCAO 래트 뇌 조직의 뉴런 세포자멸사(스케일 바:50μm); d miR-326-5p를 상향 조절하거나 STAT3를 하향 조절한 후 MCAO 쥐의 뇌 조직에서 TUNEL 양성 비율; 이 miR-326-5p를 상향 조절하거나 STAT3를 하향 조절한 후 MCAO 쥐의 뇌 조직에서 Bcl-2 및 Bax mRNA 발현; *피 가짜 그룹과 비교하여 <0.05; #피 NC 그룹과 비교하여 <0.05; &피 <0.05 miR-326-5p agomir 그룹과 비교
TUNEL 염색은 뇌 조직에서 Bcl-2 및 Bax mRNA 발현에 대한 신경 세포 사멸 및 RT-qPCR의 검출에 사용되었습니다. 결과는 (그림 3c-e) sham 그룹 및 miR-326-5p agomir 그룹과 비교하여 TUNEL 양성 비율이 증가하고 Bcl-2 mRNA 발현이 감소하고 Bax mRNA 발현이 MCAO 그룹에서 증가하는 것으로 나타났습니다. miR-326-5p agomir + oe-STAT3 그룹(모든 P <0.05). miR-326-5p agomir와 sh-STAT3 그룹에서 NC 그룹보다 TUNEL 양성률과 Bax mRNA 발현이 낮았고 Bcl-2 mRNA 발현이 높게 나타났다(모든 P <0.05).
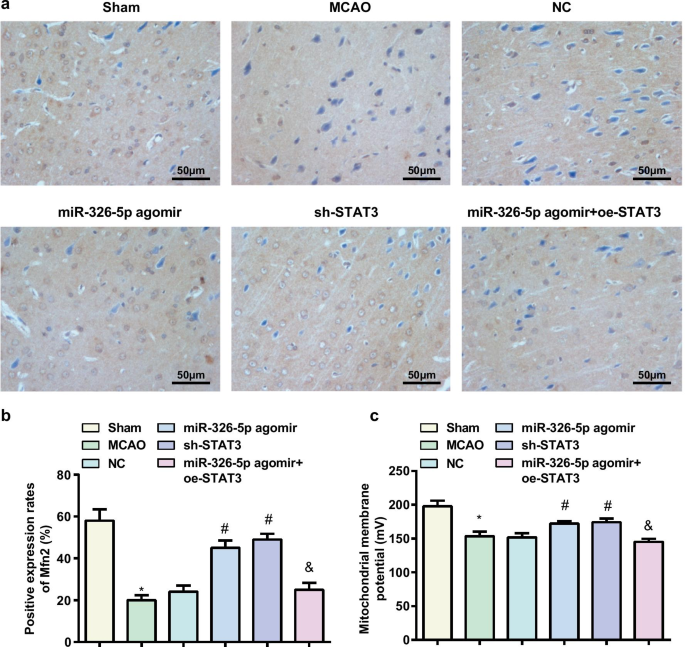
miR-326-5p의 상향 조절 또는 STAT3의 하향 조절은 CI/RI가 있는 쥐의 뇌 조직에서 미토콘드리아 막 전위 수준을 증가시킵니다.
랫트 뇌 조직에서 Mfn2 단백질 발현 검출은 면역조직화학법으로 수행되었으며, 그 결과 (그림 4a, b) 더 낮은 Mfn2 단백질 발현이 MCAO 그룹과 비교하여 sham 그룹과 miR-326-5p agomir 그룹에서 테스트되었음을 보여주었습니다. miR-326-5p agomir + oe-STAT3 그룹에 비해 Mfn2 단백질 발현은 NC 그룹 대신 miR-326-5p agomir 그룹과 sh-STAT3 그룹에서 더 높게 측정되었습니다(모든 P <0.05).
<그림>
miR-326-5p의 상향 조절 또는 STAT3의 하향 조절은 CI/RI가 있는 쥐의 뇌 조직에서 미토콘드리아 막 전위 수준을 증가시킵니다. 아 래트 뇌 조직에서 면역조직화학에 의해 검출된 Mfn2 발현(스케일 바:50μm); ㄴ miR-326-5p를 상향 조절하거나 STAT3를 하향 조절한 후 MCAO 쥐의 뇌 조직에서 Mfn2 양성 세포의 수; ㄷ miR-326-5p를 상향 조절하거나 STAT3를 하향 조절한 후 MCAO 쥐 뇌 조직의 뉴런에서 미토콘드리아 막횡단 전위; *피 가짜 그룹과 비교하여 <0.05; #피 NC 그룹과 비교하여 <0.05; &피 <0.05 miR-326-5p agomir 그룹과 비교
미토콘드리아 막 전위 수준의 검출은 (그림 4c) sham 그룹 및 miR-326-5p agomir 그룹과 비교하여 MCAO 그룹과 miR-326-5p agomir + oe-STAT3에서 미토콘드리아 막 전위 수준이 감소함을 발견했습니다. NC 그룹과 대조적으로 miR-326-5p agomir 그룹과 sh-STAT3 그룹에서 증가했지만 (모두 P <0.05).
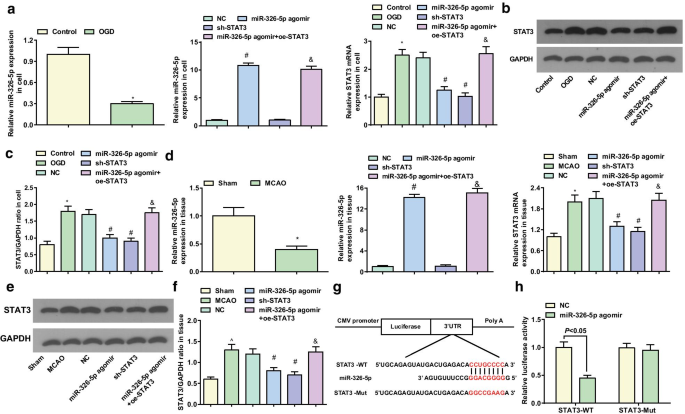
miR-326-5p 대상 STAT3
miR-326-5p와 STAT3 발현은 피질 뉴런과 뇌 조직에서 RT-qPCR과 western blot 분석을 통해 확인되었습니다. 시험관 내 피질 뉴런에서 대조군에 비해 miR-326-5p 발현의 감소와 OGD 그룹의 STAT3 발현 증가를 시사했습니다(둘 다 P <0.05). NC 그룹과 관련하여 miR-326-5p 아고미르 그룹에서 miR-326-5p 발현이 증가하고 STAT3 발현이 감소했습니다(둘 모두 P <0.05); STAT3 발현 감소(P <0.05) sh-STAT3 그룹에서. miR-326-5p agomir 그룹과 대조적으로 STAT3 발현은 miR-326-5p agomir + oe-STAT3 그룹에서 증가했습니다(P <0.05) (그림 5a-c).
<그림>
miR-326-5p는 STAT3를 표적으로 합니다. 아 시험관 내에서 OGD 처리된 피질 뉴런에서의 miR-326-5p 및 STAT3 mRNA 발현; ㄴ , ㄷ 시험관내 OGD 처리된 피질 뉴런에서의 STAT3 단백질 발현; d MCAO 쥐의 뇌 조직에서 miR-326-5p 및 STAT3 mRNA 발현; 이 , f MCAO 쥐의 뇌 조직에서 STAT3 단백질 발현; 지 생물정보학 소프트웨어에 의한 miR-326-5p 및 STAT3의 예상 결합 부위; 아 이중 루시페라제 리포터 유전자 분석에 의해 검증된 miR-326-5p와 STAT3 간의 표적화 관계; 그림 a에서 –ㄷ , *피 대조군과 비교하여 <0.05; #피 NC 그룹과 비교하여 <0.05; &피 miR-326-5p agomir 그룹과 비교하여 <0.05. 그림 d에서 –f , *피 가짜 그룹과 비교하여 <0.05; #피 NC 그룹과 비교하여 <0.05; &피 <0.05 miR-326-5p agomir 그룹과 비교
생체 내 뇌 조직에서 sham 그룹과 관련하여 MCAO 그룹에서 더 낮은 miR-326-5p와 더 높은 STAT3가 나타났습니다(둘 다 P <0.05). NC 그룹에 비해 miR-326-5p 발현은 증가하고 STAT3 발현은 miR-326-5p agomir 그룹에서 감소했습니다(둘 다 P <0.05). sh-STAT3 그룹에서 STAT3 발현 감소(P <0.05). 대조적으로 miR-326-5p agomir 그룹에서는 STAT3 발현이 miR-326-5p agomir + oe-STAT3 그룹에서 상승했습니다(P <0.05) (그림 5d–f).
STAT3 3'UTR에서 잠재적인 miR-326-5p 표적화 사이트는 DIANA 및 miRDB 소프트웨어에 의해 발견되었습니다(그림 5g). WT miR-326-5p 결합 부위와 Mut 결합 부위를 포함하는 STAT3 3'UTR을 구축하고 pmirGLO 플라스미드에 삽입했습니다. STAT3 3'UTR에 대한 miR-326-5p의 결합은 루시퍼라제 리포터 분석에 의해 추가로 검증되었습니다. 루시퍼라제 리포터 분석은 miR-326-5p 아고미르가 STAT3-WT의 상대적 루시페라제 활성을 감소시켰지만 STAT3-Mut은 감소시키지 않았음을 보여주었고(그림 5h), 이는 STAT3가 miR-326-5p의 직접적인 표적 유전자임을 암시합니다.
토론
CI/RI는 뇌혈관 사망의 주요 원인이다[22]. miRNA는 CI/RI에 관여하는 것으로 나타납니다[23]. 그러나 miR-326-5p 관련 메커니즘에 대한 포괄적인 이해는 CI/RI에서 아직 충분하지 않습니다. 따라서 이 연구는 miR-326-5p와 STAT3의 상호 결합에 중점을 두고 CI/RI에서 miR-326-5p의 구체적인 역할을 밝히는 것을 목표로 합니다. 생산적으로, 이 연구는 miR-326-5p의 상향 조절이 STAT3 억제를 통해 CI/RI를 약화시키고 Mfn2 발현을 증가시킨다고 선언했습니다.
처음에 miR-326-5p 발현은 생체 내 및 시험관 내에서 CI/RI에서 감소하는 것으로 결정되었습니다. 결과적으로, 우리는 일련의 분석을 배열하고 miR-326-5p를 상향 조절하면 뉴런 생존력과 미토콘드리아 기능이 향상되고 Mfn2 발현이 증가하며 산화 스트레스와 세포 사멸이 억제된다는 것을 발견했습니다. Furthermore, in vivo experiments in rats further verified the functional roles of restored miR-326-5p in CI/RI. As a matter of fact, high miR-326 is proved to correlate with long overall survival of patients with glioblastoma, the common type of brain tumor [24]. Also, miR-326 expression has been further evidenced to reduce in Parkinson's disease and miR-326 could suppress apoptosis of dopaminergic neurons and reduce inflammatory response [25]. Except for that, the reduction in miR-326-5p is manifested in endothelial progenitor cells in the course of myocardial infarction, and introduction of endothelial progenitor cells overexpressing could promote cardiac function recovery [11]. There has been a study illustrating that up-regulation of miR-326 improves the behavioral function, enhances neuronal viability and depresses neuronal apoptosis in mice with Alzheimer's disease [26]. Furthermore, it is documented that up-regulation of miR-326 by miR-326 mimic could suppress inducible nitric oxide synthase in dopaminergic neurons in Parkinson's disease [27].
In this study, we found that STAT3 was a target gene of miR-326-5p. Supported by an advanced study, it is indicated that STAT3 expression is negatively regulated by miR-326 in human endometrial carcinoma stem cells [28]. In the present study, we measured that STAT3 expression was enhanced in CI/RI. Currently, it has been found that I/R injury and OGD would induce STAT3 expression to increase [29]. Intriguingly, STAT3 mRNA expression trends to an increment in I/R animals [30]. Additionally, STAT3 protein expression is reported to up-regulate in CI/RI [31]. Next, our study revealed that down-regulating STAT3 had the therapeutic effects on CI/RI rats and OGD-treated neurons. As supported by a study, it is concluded that miR-31 induction discourages oxidative stress by inactivation of JAK/STAT3 pathway in ischemic stroke [32]. Also, inhibition of JAK2/STAT3 pathway is beneficial to oxidative stress impairment in CI/RI [31]. Furthermore, knockdown of JAK/STAT3 pathway is identified to enhance cell viability, and reduce oxidative stress and neuron apoptosis in ischemic brain injury [33]. Lately, inhibited JAK/STAT3 signaling pathway is witnessed to relieve myocardial infarction in myocardial I/R injury [34]. As demonstrated, depleted STAT3 undermines neuronal apoptosis in rats with white matter injury [35]. Moreover, the suppression of neuronal apoptosis, and alleviation of cerebral infarct size are attributed to JAK2/STAT3 inhibition in rats with CI/RI [36].
Finally, we focused on the effects of miR-326-5p and STAT3 on Mfn2 expression and discovered that the reduced level Mfn2 in rats with CI/RI and OGD-treated neurons could be elevated by either restoring miR-326-5p or silencing STAT3. As implicated by a previous study, it is documented that Mfn2 is decreased in the lately phase of reperfusion and poorly expressed of Mfn2 exacerbates CI/RI via restrained autophagosome formation and autophagosome and lysosome fusion [18]. Additionally, Mfn2 is implied to reduce even disappear in I/R injury in old hepatocytes [37]. However, few researches have discussed the regulatory mechanism of miR-326-5p and STAT3 for Mfn2.
Conclusion
In general, this study has elucidated the mechanisms that elevated miR-326-5p inhibits neuronal apoptosis, attenuates pathological damage of neurons and increases the expression of Mfn2 via STAT3 downregulation in CI/RI. The study may update the potential mechanism of miR-326-5p and STAT3 in CI/RI. However, more studies are still needed for further development of the molecular mechanism in CI/RI.
데이터 및 자료의 가용성
해당 없음.
약어
miRNAs:
MicroRNAs
STAT3:
Signal transducer and activator of transcription-3