이 연구에서는 좁은 크기 분포를 가진 녹색 형광 단백질(GFP) 도핑된 실리카 나노입자의 제조를 위한 새로운 접근 방식을 제시합니다. GFP는 자가형광으로 인해 모델 단백질로 선택되었습니다. 단백질이 도핑된 나노입자는 세포 내 단백질 전달 분야에서 높은 응용 가능성을 가지고 있습니다. 또한, 형광 표지된 입자를 바이오 이미징에 사용할 수 있습니다. 이러한 단백질이 도핑된 나노입자의 크기는 입자 코어 합성에 이어 쉘 재성장 단계를 포함하는 다단계 합성 공정을 사용하여 15에서 35nm로 조정되었습니다. GFP는 원 포트 반응에 의해 코어나 쉘 또는 둘 모두의 실리카 매트릭스에 선택적으로 통합되었습니다. 수득된 나노입자는 입자 크기, 유체역학적 직경, ζ-전위, 형광 및 양자 수율의 결정에 의해 특성화되었다. 측정은 GFP의 형광이 입자 합성 동안 유지되었음을 보여주었다. 세포 흡수 실험은 GFP가 도핑된 나노입자가 안정적이고 효과적인 형광 프로브로 사용될 수 있음을 보여주었습니다. 이 연구는 기능적 생물학적 거대분자를 실리카 나노입자에 통합하기 위해 선택된 접근 방식의 잠재력을 보여줌으로써 세포 내 단백질 전달과 같은 새로운 응용 분야를 열어줍니다.
<섹션 데이터-제목="배경">
배경
최근 몇 년 동안, 단백질을 마이크로 및 나노 입자로 캡슐화하는 것은 바이오센서[1] 또는 바이오리액터[2]와 같은 물질의 광범위한 응용 가능성으로 인해 널리 주목받고 있으며, 더 나아가 제어된 단백질 전달 분야[3], 세포 내 단백질 전달 [4] 및 조직 공학 [5]. 이러한 많은 응용 분야에서 캡슐화된 효소의 촉매 활성은 이러한 물질의 기본 기능 중 하나입니다. 대조적으로, 그러한 나노물질의 잠재적 화물로서 약학적으로 관련된 단백질, 펩타이드 호르몬 또는 항체는 조직 또는 세포 내 표적의 특이적 결합에 의해 기능을 발휘한다. 따라서 이러한 모든 응용 프로그램의 전제 조건 중 하나는 화물 단백질의 완전한 형태와 기능을 유지하는 것입니다. 나노구조 시스템은 작은 크기, 큰 비표면적 및 기타 고유한 특성으로 인해 생물의학 연구에서 가장 빠르게 발전하는 분야 중 하나가 되었습니다[6]. 따라서 설계된 시스템의 기능과 안정성을 개선하기 위한 새로운 미립자 담체의 개발은 이 분야에서 중요한 주제입니다[7]. 나노입자 담체의 매트릭스는 탄수화물, 지질 또는 중합체와 같은 생체거대분자 또는 유기 성분을 기반으로 하여 고체-지질 나노입자, 리포솜 또는 덴드리머와 같은 시스템을 형성할 수 있습니다. 또한, 나노구조 시스템은 금속이나 산화물과 같은 무기 물질을 기반으로 할 수도 있습니다[8]. 이러한 모든 재료 시스템은 다양한 공통 및 특정 요구 사항을 충족해야 합니다. 우선, 매트릭스 재료는 안전한 적용을 용이하게 하기 위해 생체 적합성이어야 합니다[9]. 둘째, 시스템의 수명 주기 동안 캐리어 재료로서의 기능을 수행할 수 있을 만큼 충분히 안정적이어야 합니다. 또한 상당한 단백질 부하 및 보유 능력과 단백질 방출 조절 능력을 제공해야 합니다[10].
흡착 또는 공유 결합을 통해 나노 물체의 표면에 단백질이 부착되는 것 외에도[11], 단백질은 나노 구조 내에 포획되어 안정성과 효소 활성을 향상시킬 수 있습니다[2]. 나노포집은 졸-겔 공정[12] 또는 유중수 마이크로에멀젼 접근법을 통해 실리카 전구체의 가수분해 및 축합에 의해 달성될 수 있으며, 이는 물-오일 계면에서 쉘을 둘러싼 효소의 중합을 유발합니다[13]. 이러한 방법에서 단백질의 포획은 공유 또는 비공유 결합 과정을 사용하는 두 가지 다른 화학적 접근에 의해 발생할 수 있습니다[14]. 특히, 비정질 이산화규소는 높은 생체적합성, 불활성 및 기계적 안정성으로 인해 단백질의 유망한 담체 물질이다[15]. 다양한 경로, 특히 이산화규소로의 효소 캡슐화를 위한 생체모방 접근법이 따랐으며[2, 16], 효소의 방출 프로파일은 링커의 화학 반응 또는 실리카 매트릭스의 분해에 의해 제어됩니다. 메조포러스 물질은 또한 2-50nm의 기공 내에서 효소를 고정하기 위한 매트릭스로 사용되었습니다[13, 17]. 메조포러스 나노입자로부터의 화물 방출은 "게이트키퍼" 전략을 사용하거나 약물과의 결합 친화성을 제어하기 위해 기공의 내부 표면을 수정하여 조정할 수 있습니다[10b]. 그럼에도 불구하고, 기공 크기는 수행된 메조포러스 실리카 스캐폴드로의 효소 로딩을 제한할 수 있으며 [18], 이것이 단백질 전달에 대한 새로운 전략이 최근 조사 중인 이유입니다.
실리카 나노입자는 바이오이미징에 널리 사용되기 때문에[19], 형광 단백질의 통합은 생체적합성 형광 프로브의 생성을 위한 하나의 옵션을 구성합니다. 예를 들어, 역 에멀젼 기술을 통해 실리카 나노입자에 녹색 형광 단백질(GFP)을 통합하는 것은 문헌[20]에 설명되어 있습니다. 이러한 연구는 GFP를 실리카 입자 매트릭스에 통합하면 단백질의 형광 강도뿐만 아니라 열 안정성, 화학적 변성 및 프로테아제 처리에 대한 안정성을 향상시킬 수 있음을 나타냅니다. 그럼에도 불구하고, 이 방법은 좁은 크기 분포를 가진 더 낮은 나노규모 범위에서 잘 정의된 실리카 나노입자의 합성에 덜 적합합니다. 또한 합성 조건에는 계면활성제, 알코올 또는 고알칼리성 염기에 대한 접촉과 고온이 포함되어 있어 모두 민감한 단백질의 통합에 적합하지 않을 수 있습니다[20, 21].
따라서 우리는 GFP를 모델 단백질로 사용하여 단백질 도핑된 실리카 나노입자의 제조를 위한 새로운 접근 방식에 대해 보고합니다. 이를 위해 우리는 순한 합성 조건(실온, 낮은 염도)에서 원 포트 합성을 사용한 후 정제를 위한 투석 단계를 사용했습니다. 이 접근 방식은 50nm 미만의 크기 영역에서 좁은 크기 분포를 나타내는 단백질이 포함된 실리카 나노입자를 제조할 수 있는 가능성이 특징입니다.
방법
자료
모든 화학 물질은 Sigma-Aldrich(독일 Taufkirchen)에서 구입한 대로 추가 정제 없이 사용했습니다. 모든 합성 및 정제 단계에서 초순수(18.2MΩ, Milli-Q 정수 시스템 type ELIX 20, Millipore Corp., USA)를 사용했습니다.
GFP의 준비
GFP는 다른 곳에서 설명한 대로 단백질 발현 및 후속 정제에 의해 얻어졌습니다[22]. 요약하면, N-말단 His6-tag를 포함하는 GFP는 E. 대장균 XL1-Blue 및 Ni-하전 친화성 크로마토그래피(Qiagen, Hilden, Germany)에 의해 정제되었습니다. 이어서, 완충제 교환을 위해 단백질을 농축기 장치(3 kDa 분자량 차단(MWCO) 멤브레인, Pall, Dreieich, Germany)로 옮겼다. GFP는 각각 15mL의 ʟ-아르기닌 및 중탄산나트륨 용액을 첨가하여 3회 세척한 다음, 3mL의 ʟ-아르기닌/중탄산나트륨 용액에서 회수했습니다. 그 후, GFP-현탁액을 멸균 0.22μm 셀룰로오스 아세테이트 필터(Carl Roth, Karlsruhe, Germany)를 통해 멸균 튜브로 여과했습니다. 사용하기 전에 단백질 농도를 1mg mL
−1
로 조정했습니다. 7.2mmol L
−1
ʟ-아르기닌(pH =10.3) 또는 10.0mmol L
−1
NaHCO3 (pH =9.2) 용액.
나노 입자의 합성 및 정제
실리카 나노입자는 이전에 설명된 수정된 프로토콜에 따라 제조되었습니다[23]. 간단히 말해서, 비극성 전구체로 사용되는 테트라에톡시실란(TEOS)은 ʟ-아르기닌 촉매 작용에 의해 매개되는 2상 물/시클로헥산 시스템에서 가수분해되었습니다.
핵심 입자의 준비
3구 둥근 바닥 플라스크에서 91mg(0.52mmol) ʟ-아르기닌을 69mL의 물에 용해시킨 후 4.5mL의 사이클로헥산을 상부 층으로 첨가했습니다. 반응 혼합물을 교반 하에 40℃로 가열하였다. 5.5mL(24.63mmol) TEOS를 첨가한 후 혼합물을 이러한 조건에서 추가로 20시간 동안 유지했습니다.
실리카 쉘 레이어
후속 쉘 성장 단계의 경우 코어 입자 또는 첫 번째 쉘 성장 단계에서 생성된 입자가 사용되었습니다. 껍질 성장을 위해 14mg(0.08mmol) ʟ-아르기닌을 물 36mL에 용해시키고 미리 준비한 입자 분산액 10mL를 첨가했습니다. 5mL의 사이클로헥산을 첨가한 후, 혼합물을 40°C로 가열했습니다. 3.52mL(15.8mmol) TEOS를 첨가한 후, 혼합물을 추가로 20시간 동안 교반했습니다.
GFP 도핑된 나노입자의 준비. GFP 도핑된 나노입자의 제조를 위해 TEOS를 첨가한 지 30분 후에 200μg(6.9nmol)의 GFP가 첨가되었습니다.
입자 정화
나노입자는 셀룰로오스 수화물 멤브레인(Nadir-dialysis tubing, MWCO 10kDa, Carl Roth, Karlsruhe, Germany)을 사용하여 4시간 동안 물(4L, 30, 90, 180분 후 물 교환)에 대한 후속 투석으로 정제했습니다. 마지막으로, 나노입자는 멸균 0.22 µm 셀룰로오스 아세테이트 멤브레인 필터(Carl Roth, Karlsruhe, Germany)를 사용하여 멸균 플라스크로 여과되었습니다.
투과 전자 현미경(TEM)
형태 및 평균 입자 직경은 JEM-2100F 현미경(JEOL, Freising, Germany)을 사용하여 결정하였다. 입자 크기 분포는 X-ImageJ 소프트웨어(버전:1.45 s, National Institute of Health(http://rsb.info.nih.gov/ij)의 winPenPack X-ImageJ Launcher)를 사용하여 50개 나노입자의 무작위 샘플에서 결정되었습니다. /).
유체역학적 직경
나노입자의 유체역학적 직경은 Zetasizer Nano ZSP(Malvern Instruments, Herrenberg, Germany)를 사용하여 기록되었습니다. 측정 전에 입자 분산액을 물에 1:10 희석했습니다. 측정은 25°C에서 수행되었습니다. 각 샘플은 3 × 15번 측정되었습니다. 직경은 부피 분포의 계산에 의해 결정되었습니다. 이것은 Mie 이론을 사용하여 강도 크기 분포에서 변환되었습니다.
ζ 가능성
ζ 전위는 샘플을 0.01M KCl(9:1)에 희석한 것을 제외하고는 동일한 장비를 사용하여 위에서 설명한 조건으로 측정되었습니다.
분석용 초원심분리(AUC)
침강 속도를 측정하기 위해 AnTi60 로터가 있는 수정된 Beckman-Coulter XL-80 K. 실험을 위해 온도를 20°C로 설정하고 속도를 10,000rpm으로 설정하고 21번의 스캔을 수행했습니다. 파장은 실리카의 경우 261nm, GFP 감지의 경우 488nm로 설정되었습니다.
형광 분광기
Fluoromax-3 분광형광계(Spex, Horiba Scientific, Oberursel, Germany)를 사용하여 나노입자, 순수 GFP 및 침출 실험의 여과액의 형광 스펙트럼을 기록했습니다. 측정을 위해 순수한 GFP, 입자 분산액 및 여액을 물에 1:10 희석했습니다. 여기 파장은 488nm로 설정되었으며 스펙트럼은 498~800nm의 스펙트럼 범위에서 기록되었습니다.
형광 양자 수율
얻어진 나노 입자 및 순수한 GFP의 양자 수율은 Williamson et al.의 상대 방법을 사용하여 결정되었습니다. [24]. GFP에 대한 기준으로 로다민 6G 및 Atto488을 사용했습니다. 기준 염료와 혼합된 도핑되지 않은 나노입자를 사용하여 비교 측정을 수행했습니다. 450nm의 여기 파장을 사용하여 형광 스펙트럼을 기록했습니다. 추가 UV/vis 측정은 Varian Cary 300 Scan UV(Agilent Technologies, Darmstadt, Germany)를 사용하여 수행되었습니다.
여기서 φP 제품의 양자 수율, φS 참조의 양자 수율. 용어 기울기S 및 기울기P 각각 참조 및 제품의 흡광도 대 통합 형광 강도의 플롯에서 파생된 기울기를 나타냅니다. nP 그리고 nS 사용된 용매의 굴절률에 해당합니다[25].
단백질 누출
침출 실험을 위해 희석되지 않은 입자 분산액을 원심분리(16,000g, 5분)에 의해 변형된 폴리에테르 설폰 멤브레인(MWCO =100kDa 또는 300kDa, Pall, Dreieich, Germany)을 통해 한외여과했습니다.
열 안정성
열 안정성 분석을 위해 나노입자와 순수한 GFP를 20 또는 60°C에서 0시간과 24시간 동안 보관했습니다. 나노 입자와 순수한 GFP는 위에서 설명한 대로 희석되었습니다.
광표백
광표백에 대한 GFP 도핑된 나노입자 및 순수 GFP의 안정성을 조사하기 위해 용액을 최대 20분의 기간 동안 7개의 녹색 LED에서 방출되는 빛에 노출시켰습니다. t에서 채취한 샘플의 형광 강도 =0, 2, 20분이 측정되었습니다.
단백질 분해에 대한 안정성
단백질분해효소 K에 대한 GFP의 안정성을 결정하기 위해 순수한 GFP, 표지되지 않은 실리카 나노입자(CU S1U S2U ) 추가 GFP 및 3회 표지된 실리카 나노입자(CF S1F S2F )는 동일한 GFP 농도 및 동일한 양의 입자로 사용되었습니다. 모든 샘플은 1:100으로 희석되었습니다. 10개의 GFP 분자의 양에 대해 하나의 프로테이나제 K 분자가 선택되었습니다. 효소를 첨가하기 전에 위의 조건으로 하나의 샘플을 측정하였다. 추가 후 측정은 t 후에 수행되었습니다. =0, 15, 30, 45, 60, 90분
세포 흡수 실험
세포에 의한 나노입자 및 GFP의 내재화를 결정하기 위해 폐암 세포주 A549(ACC-107)를 사용하여 세포 흡수 실험을 수행했습니다.
세포 배양
A549-세포(DSMZ, Braunschweig, Germany)는 10% 태아를 함유하는 Dulbecco's Modified Eagle Medium(DMEM, Thermo-Fisher-Scientific, Waltham, MA, USA)을 사용하여 T75-플라스크(Greiner bio-one, Frickenhausen, Germany)에서 배양되었습니다. 송아지 혈청(FCS). 2 × 10
4
cm
−2
A549 세포를 12웰 플레이트의 커버 슬립에 접종하고 24시간 동안 배양했습니다. 그런 다음 세포를 24시간 동안 1mL 배지에서 GFP 도핑된 나노입자 및 GFP 용액으로 처리했습니다. SiO2 나노입자의 농도는 37 μg mL
−1
였습니다. GFP 농도는 5μg mL
−1
였습니다. 나노 입자와 순수한 GFP 모두에 대해. 처리 후 세포를 인산완충식염수(PBS)로 두 번 세척했습니다.
샘플 준비 및 공초점 이미징
세포를 실온에서 20분 동안 PBS 중 4% 파라포름알데히드로 고정했습니다. 세포막 염색의 경우, 테트라메틸로다민-접합 WGA(밀배아 응집소(2 μg mL
−1
(PBS에서), W849, Thermo-Fisher-Scientific(Invitrogen), Waltham, MA, USA)를 첨가하고 실온에서 10분 동안 인큐베이션했습니다. PBS로 3단계 세척한 후, 세포를 PBS로 3회 세척하고 Mowiol/DABCO(Carl Roth, Karlsruhe, Germany)를 사용하여 유리 슬라이드에 장착했습니다.
공초점 이미지는 TCS SP5 시스템(Leica, Wetzlar, Germany)에서 촬영되었습니다. 이미징의 경우 63배 오일 침수 목표(n =1.518)이 사용되었습니다. λ에서 아르곤 이온 레이저 라인을 사용하여 순차 스캔을 수행했습니다. =GFP의 여기를 위한 488nm(25%) 및 λ에서 다이오드 펌핑 고체 상태 레이저 =테트라메틸로다민의 여기를 위한 561 nm(25%).
결과 및 토론
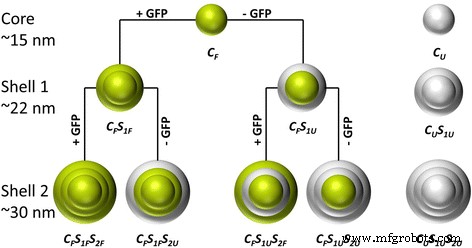
이 연구는 단백질의 생화학적 특성과 기능을 유지하는 적절한 조건에서 GFP로 실리카 나노입자를 기능화하는 것을 목표로 합니다. 이전 연구에서 우리는 2상 시클로헥산/물 시스템에서 테트라에톡시실란(TEOS)의 ʟ-아르기닌 제어 가수분해를 사용하여 15~80nm 크기 범위의 근적외선 염료 도핑된 단분산 형광 실리카 나노입자를 합성했습니다[26]. 여기에서 우리는 GFP를 모델 단백질로 실리카 매트릭스에 삽입하기 위해 이 합성 절차를 채택했습니다. 반응식 1에서 입자 합성 절차가 개략적으로 도시되어 있습니다. GFP 도핑 및 비도핑 구조(코어/쉘)는 각각 녹색과 회색으로 강조 표시됩니다. 첫 번째 단계에서 GFP가 도핑된 실리카 코어 입자(CF ) 얻었다. 후속 재성장 단계(CF S1 및 CF S1 S2 ) 더 큰 입자 크기의 합성을 허용했습니다. 첫 번째 재성장 단계에서 쉘은 (CF S1F ) 또는 미포함(CF S1U ) 단백질의 혼입. 유사하게, 두 번째 재성장 단계에서 레이블이 지정된 (CF S1F S2F , CF S1U S2F ) 또는 레이블이 없는(CF S1F S2U , CF S1U S2U ) 쉘이 추가되었습니다. 이러한 변형을 통해 포함된 단백질의 양과 지정된 껍질 또는 입자 코어로의 맞춤 배열을 훌륭하게 제어할 수 있습니다. 또한 GFP가 포함되지 않은 순수 실리카 나노입자(CU , CU S1U , 및 CU S1U S2U ) 입자 특성에 대한 단백질 임베딩의 잠재적 영향을 조사하기 위해 합성되었습니다. 또한, 이 모든 단계에서 GFP는 두 가지 다른 완충 시스템(ʟ-아르기닌 및 NaHCO3 ) 다양한 pH 값의 입자 합성, 형태, 형광 강도, 방출 파장 및 ζ 전위에 대한 단백질 용매의 영향을 결정합니다.
<그림>
합성된 입자 및 입자 구조의 개요. 녹색은 각각 코어 또는 쉘에 GFP가 포함되었음을 나타냅니다. 회색은 GFP가 없는 껍질을 나타냅니다(CF =코어 형광등, CU =레이블이 없는 코어, SF =쉘 형광등, SU =라벨이 없는 쉘, S1 =첫 번째 셸 레이어, S2 =두 번째 셸 레이어)
나노입자 특성화
물리적 입자 속성의 결정
GFP 통합 후 입자 크기와 형태를 설명하고 이러한 특성에 대한 두 가지 다른 완충 시스템의 영향을 결정하기 위해 TEM 이미지를 기록했습니다(그림 1). GFP(NaHCO3의 추가 TEM 이미지 ) 수정, GFP(ʟ-아르기닌) 수정 및 레이블이 지정되지 않은 나노 입자는 SI에 표시됩니다(추가 파일 1:그림 S1, 추가 파일 2:그림 S2, 추가 파일 3:그림 S3, 추가 파일 4:그림 S4). 두 개의 재성장 단계를 통한 합성 절차에 따라 세 가지 다른 입자 크기가 얻어졌습니다. 코어 입자의 크기는 약 15nm, 첫 번째 재성장 단계 후의 입자는 약 22nm, 두 번째 단계 이후의 입자는 약 32nm였습니다. 요약하면, 모든 나노입자는 거의 구형이었고 좁은 크기 분포를 나타냈습니다(p <10%). 완전히 염색된 3세대 GFP(ʟ-아르기닌) 나노 입자(CF , CF S1F , 및 CF S1F S2F ) 및 GFP(NaHCO3 ) (CF ) 코어 나노입자를 모델로 선택했습니다.
<그림>
3세대 GFP-ʟ-아르기닌 변형 나노입자와 GFP(NaHCO3 코어 입자)의 TEM 이미지 ) 변형된 나노입자. a에서 , ㄷ 그리고 d , GFP(ʟ-아르기닌)의 3세대가 표시됩니다:CF 코어 입자(a , dTEM =15.5 ± 1.1 nm), CF S1F 첫 번째 재성장 단계 후 나노 입자(코어 + 쉘 1) (c , dTEM =23.5 ± 2.0 nm) 및 CF S1F S2F 두 번째 재성장 단계 후(코어 + 쉘 1 + 쉘 2) (d , dTEM =35.3 ± 2.0 nm). b에서 , GFP(NaHCO3 )-표지된 코어 나노입자(dTEM =15.2 ± 1.2 nm) 표시됨
상이한 GFP 도핑 및 표지되지 않은 나노입자의 크기를 비교하면(표 1), 동일한 수의 재성장 단계가 단백질의 존재 또는 사용된 완충 용액과 무관하게 동일한 평균 입자 크기를 초래한다는 점은 주목할 만하다. 레이블이 지정되지 않은 입자도 비슷한 크기를 가졌습니다(CU :dTEM =13.4 ± 0.4nm, dDLS =10 ± 3 nm; CU S1U :dTEM =20.9 ± 1.3nm, dDLS =20 ± 6 nm; CU S1U S2U :dTEM =33.2 ± 1.0nm, dDLS =38 ± 10nm).
결론적으로, 단백질이 제공된 실리카 기질과 완충용액에 단백질의 혼입은 생성된 입자 크기 및 형태에 큰 영향을 미치지 않음을 입증하였다.
우리가 아는 한, 유사한 작은 크기와 똑같이 좁은 크기 분포(<10%)를 나타내는 문헌에 기술된 다른 GFP 내장 실리카 나노입자는 없습니다[20, 27]. 이러한 작은 나노입자는 암 진단 및 치료뿐만 아니라 세포 내 단백질 전달 분야에서 유망한 응용 가능성을 가지고 있습니다[28].
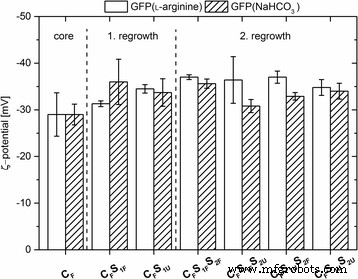
ζ-잠재력
모든 나노 입자의 ζ 전위는 전기 영동 이동성을 사용하여 계산을 통해 결정되었습니다. 모든 유형의 도핑된 나노입자는 - 28 ~ - 36 mV 범위의 절대값으로 음의 ζ 전위를 나타냈습니다(그림 2). 이에 비해 비표지 입자의 ζ 전위는 코어 입자의 경우 - 35.5 ± 2.0mV, 첫 번째 재성장 단계 후 - 34.0 ± 3.7mV 및 두 번째 단계 후 - 34.5 ± mV1.2로 매우 유사한 값을 나타냅니다. 이러한 높은 음의 ζ 전위(<− 28mV) 값은 정전기 반발로 인한 덩어리에 대한 나노 입자의 높은 안정성을 나타냅니다. 표지되지 않은 나노입자의 ζ 전위와 비교하여 데이터는 생성된 입자 크기나 입자 코어 또는 쉘의 입자 매트릭스에 GFP를 통합하는 것이 입자 전하에 상당한 영향을 미치지 않는다는 것을 나타냅니다.
<그림>
표지된 나노입자의 ζ-전위 [mV]. 나노입자는 7.2mM ʟ-아르기닌 또는 10mM NaHCO3에 용해된 GFP에서 시작하여 준비되었습니다. . 오차 막대 세 가지 측정에서 파생된 표준 편차를 나타냅니다.
분광학 연구
형광 분광기
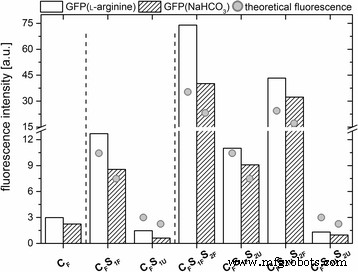
모든 GFP 도핑된 실리카 나노입자는 유사한 최대 방출(λ그들 =508 nm), 이는 유리 GFP의 최대 방출과도 비슷했습니다(SI, 추가 파일 5:그림 S5). 다양한 표지된 나노입자의 형광 강도를 비교하기 위해 나노입자 농도를 정규화했습니다(SI 5의 계산). 예상대로, 표지된 쉘을 단계적으로 추가하면 나노입자의 형광이 증가합니다(그림 3).
<그림>
다양한 입자 시스템 각각에 대해 508nm에서 최대 방출의 정규화된 형광 강도입니다. 또한 이론적인 형광 강도(회색 점 ) 입자 부피의 증가와 관련하여 입자의 크기가 표시됩니다.
표지된 코어만 있고 도핑되지 않은 껍질이 있는 나노입자는 가장 낮은 형광성을 나타냈습니다. 하나의 추가로 표지된 껍질을 가진 나노입자는 중간 형광을 보였고, 두 개의 표지된 껍질을 가진 나노입자는 가장 강한 형광을 보였다(그림 3). 놀랍게도, 도핑되지 않은 외부 쉘의 추가는 도핑된 외부 층을 갖는 나노입자와 비교하여 나노입자의 형광을 약간 감소시키는 것으로 보였다. 이 효과는 라벨이 없는 실리카 쉘의 차폐 효과에 의해 유도될 수 있습니다. 요약하면, 코어 입자에 GFP가 도핑된 쉘을 추가하면 생성된 나노입자의 형광 강도가 증가하며 이는 나노입자 성장에 수반되는 부피 변화와 상관관계가 있는 것으로 보입니다.
정제 후 초기에 ʟ-아르기닌에 용해된 GFP의 포매는 NaHCO3에 용해된 GFP에서 시작하여 아날로그 포매 공정을 통해 얻은 나노입자와 비교하여 생성된 나노입자의 형광 강도가 1.3배 더 높았습니다. . 유사하게, ʟ-아르기닌에 희석된 GFP는 NaHCO에 희석된 GFP에 비해 더 높은 형광 강도를 나타냈습니다3 (추가 파일 5:그림 S5). 이 효과는 완충액의 다양한 pH 값으로 설명될 수 있습니다(pHʟ-arginine =10.3, pH\( _{{\mathrm{NaHCO}}_3} \) =9.2).
이러한 이유로 순수한 GFP의 형광은 pH 값의 함수로 체계적으로 측정되었습니다(SI, 추가 파일 6:그림 S6). 데이터는 pH 5.5 - 10.5 범위에서 pH가 증가함에 따라 형광의 쌍곡선 모양 증가를 보여주었습니다. 결과는 GFP의 pH 의존적 형광에 대한 다른 보고서와 일치합니다. 야생형 GFP의 경우, 형광은 pH 6 - 10 범위에서 변경되지 않지만 낮은 pH에서 감소하고 pH 값> 10에서 증가하는 것으로 보고되었습니다[29]. 또한 GFP의 pH 민감도는 점 돌연변이의 도입으로 수정될 수 있습니다[30]. 본 연구에 사용된 GFP는 Aequorea와 비교하여 3점 돌연변이를 가지고 있습니다. 야생형 단백질, 즉 S2A, F64L, S65T. 이들 중 트레오닌에 대한 65번 위치의 세린 치환은 이 아미노산이 발색단의 형성에 관여하기 때문에 480nm에서 여기될 때 단백질의 형광 강도를 증가시키는 것으로 나타났습니다. 또한 S65T/F64L 변이체는 pH 의존적 형광을 나타냅니다[30]. GFP 도핑된 나노입자(CF )는 유사한 pH 의존성 형광을 나타내어(그림 3), pH 의존성 메커니즘이 임베딩 프로세스에 영향을 받지 않음을 나타냅니다.
형광 양자 수율
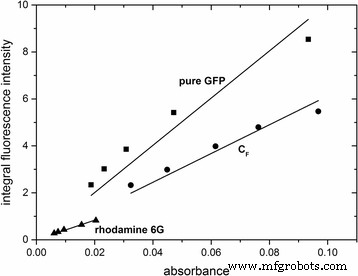
형광 나노 입자의 특성을 추가로 특성화하기 위해 양자 수율을 결정했습니다. 이는 통합된 형광 강도 대 488nm에서 흡광도를 표시하여 달성했습니다(그림 4). 그 후, 양자 수율은 수학식 1을 사용하여 계산되었다. 2. 로다민 6G를 기준으로 GFP가 도핑된 나노입자 CF의 양자 수율 S1F 순수한 GFP는 φ\( _{{\mathrm{C}}_{\mathrm{F}}{\mathrm{S}}_{1\mathrm{F}}} \) =0.62로 결정되었으며 φ순수GFP =0.38, 각각. 두 번째 참조로 Atto488을 사용하여 결과를 확인했습니다(SI, 추가 파일 7:그림 S7). 순수한 GFP와 비교하여 GFP가 도핑된 나노입자의 더 높은 양자 수율은 GFP가 실리카 매트릭스에 캡슐화되어 발생하는 것으로 보이며 단백질의 공간적 고정 또는 실리카 매트릭스에 의해 제공되는 변경된 화학적 환경과 연결될 수 있습니다. .
<그림>
GFP가 도핑된 입자와 순수 GFP의 통합 형광 강도 대 488nm에서의 흡광도 플롯입니다. 로다민 6G를 기준으로 사용했습니다. 선형 상관 관계는 직선으로 적합했습니다. . 해당 선형 방정식은 다음과 같습니다. y순수GFP =1.00554 × 10
10
× x , R
2
=0.97712; y\( _{C_F{S}_{1F}} \) =6.12332 × 10
9
× x , R
2
=0.99331; 예로다민6G =4.1772 × 10
9
× x , R
2
=0.99678
입자 안정성
단백질 누출
GFP가 도핑된 나노입자의 결합 안정성을 입증하기 위해 침출 실험을 수행하였다. GFP(MW~27kDa)의 통과를 허용하지만 나노입자를 유지하는 MWCO로 막을 통한 한외여과 후, 여과액에서 형광이 검출되지 않아 GFP가 실리카 매트릭스에 영구적으로 결합되었음을 나타냅니다.
분석용 초원심분리
얻어진 결과를 뒷받침하고 입자 매트릭스에 결합하는 GFP의 유형을 결정하기 위해 분석 초원심분리를 수행했습니다. 이를 위해 CF 레이블이 지정됨 S1F S2F 입자 및 레이블이 지정되지 않은 CU S1U S2U particles mixed with GFP were measured at the same particle and GFP concentrations. The results (Additional file 8:Figure S8 in the SI) indicate that most of the GFP molecules are embedded into the silica matrix during the synthesis.
Thermal Stability
To determine their thermal stability, the fluorescence signals of CF in comparison to pure GFP were measured after incubation at room temperature and 60 °C respectively (Fig. 5). After 24 h at room temperature, no decrease in the fluorescence of both samples was detectable, indicating no influence on the protein stability. However, after 24 h at elevated temperature of 60 °C, only 20% of the initial fluorescence intensity of CF could be observed, whereas no fluorescence signal of pure GFP reverted. This strongly indicates a higher thermal stability of GFP-embedded silica compared to pure GFP. Since an elevated temperature leads to a significant increase in the thermal motions of the protein molecule, which can disrupt its structure, it is hypothesised that the surrounding silica matrix protected the GFP against external influences by spatial constraints.
Influence of temperature (r.t., 60 °C) on the fluorescence of GFP-doped particles (CF , ʟ-arginine) and pure GFP. The normalised fluorescence intensity [%] of the emission maximum at 508 nm versus time [h] is shown
Photostability
Furthermore, the photostability of the samples was tested. For measurements, the nanoparticle stock suspension (CF , ʟ-arginine) was diluted tenfold. Pure GFP was diluted in ʟ-arginine according to the calculated concentration of GFP in the nanoparticle suspension. After exposure of the samples to light of a green LED array over a period of time up to 20 min, the fluorescence intensity was measured (Fig. 6). Within 20 min, the fluorescence intensity of the nanoparticle suspension decreased only slightly. After 20 min, 89% of the initial fluorescence (100%) of the nanoparticles was preserved. In comparison, the pure GFP seemed to be more affected by light exposure. After 20 min, only 81% of the initial fluorescence of pure GFP remained. This result indicated, that GFP, when embedded into silica nanoparticles, was better protected from photochemical alterations induced by the LED light than the pure protein.
Photostability of GFP-doped nanoparticles (CF ) and pure GFP in ʟ-arginine. The normalised fluorescence intensity [%] of the emission maximum at 508 nm was measured after exposure to LED light for the given times. Data are mean values. Error bars indicate the standard deviation
Stability Against Protein Degradation
As a further characterisation step, the degradation of GFP in the presence of proteinase K was tested. Therefore, three different systems were used (pure GFP, unlabelled CU S1U S2U mixed with GFP and labelled CF S1F S2F ). For all systems, equal amounts of GFP and particles were used. After 90 min of incubation, the fluorescence intensity of pure GFP and unlabelled particles with added GFP decreased to 5 - 7% of the initial fluorescence intensity, whereas the one of the labelled particles decreased to 52% (Fig. 7). This result indicates that the GFP is protected by the silica matrix and is degraded slower than free GFP in presence of proteolytic enzymes.
Stability against protein degradation of pure GFP (grey ), unlabelled particles mixed with GFP (CU S1U S2U , blue ), and GFP-doped silica nanoparticles (CF S1F S2F , green ). The normalised fluorescence intensity [%] of the emission maximum at 508 nm was plotted against the incubation time [min] with proteinase K
To conclude, the encapsulation of GFP into silica matrix appeared to bring about significant advantages:The stability of the protein was increased not only against elevated temperatures and light-induced photobleaching but also against the degradation through enzymes. Therefore, the silica matrix seems to protect the embedded GFP as compared to the free GFP.
Cellular Uptake Experiments
In order to determine, if the GFP-doped nanoparticles are able to deliver their cargo into cells, uptake experiments were performed (Fig. 8). A549 cells were exposed to GFP-doped nanoparticles and for comparison to the pure protein. In order to optimise the GFP load of the particles for imaging, a higher amount of GFP as compared to the nanoparticles described before was embedded into the particles. More specifically, a 20-fold amount of GFP in ʟ-arginine was used to label the second shell of the CF S1F S2F 입자. These nanoparticles were diluted to a final concentration of 37 μg SiO2 per millilitre in cell culture medium and incubated for 24 h with the cells. The amount of GFP in both samples (nanoparticles and pure GFP) was 5 μg mL
−1
.
Confocal microscopy images of A549 cells after 24 h exposure to GFP dissolved in ʟ-arginine (A1 –A3 ) and GFP-doped nanoparticles CF S1F S2F (B1 –B3 ), and control cells (C ). Top (1):merge-images; middle (2):Cell membrane (WGA):red; bottom (3):GFP, green . Arrows indicate internalised nanoparticles. Contrast and brightness were enhanced by using the ImageJ software
In order to visualise the cells, the cell membrane was labelled, using tetramethylrhodamine-coupled WGA (wheat germ agglutinin). Confocal imaging was used to localise the GFP-doped nanoparticles and the pure GFP in the cells. After exposure of cells to GFP, no signal related to GFP was observed inside the cell bodies (Fig. 8a). Compared to the control cells, no difference in signal intensity of both channels could be observed (Fig. 8c).
In contrast, after exposure of the cells to the GFP-loaded nanoparticles, bright fluorescence signals were detected in the perinuclear region, indicating internalisation of the loaded nanoparticles through endocytosis. The GFP-loaded nanoparticles appeared to be excluded from the nuclear compartment. A second fraction of agglomerated nanoparticles was detected on top of the cell membrane (Fig. 8b).
In conclusion, the GFP-doped nanoparticles are internalised by the cells and are able to transport their cargo into the cells. After exposure of the cells to GFP, fluorescence signals were not detected inside the cell body. This finding is in line with the results of Pesce et al. [31], who did not observe cell-associated fluorescence after incubation of A549 cells with GFP for 24 h. The lack of cell-associated GFP signals might be due to the fact that GFP is not internalised by the cells. Alternatively, GFP fluorescence might be quenched by the low pH value present in endocytic vesicles or lysosomes or degraded by proteolytic enzymes. Therefore, the fluorescence signals of the nanoparticles might indicate a protective effect of the silica nanoparticle matrix against lysosomal degradation.
Conclusions
In this study, a novel approach is presented for synthesis of monodisperse GFP-doped silica nanoparticles with a mean particle-core size of 15 nm. By subsequent growth steps, the particle size and the amount of embedded GFP can be varied. At the end of this procedure, the fluorescence properties of GFP are kept. Incorporation of GFP into additional outer shells results in an increase in the nanoparticle fluorescence. Coverage of the nanoparticles by non-doped shells seems to slightly decrease their fluorescence. These properties indicate the potential to incorporate cargo molecules into specific particle shells. The GFP-doped nanoparticles exhibit a higher quantum yield as compared to the pure GFP. The incorporation into the silica matrix appeared to be durable, as no leaching of protein was detected by ultrafiltration. The silica matrix also seems to improve the thermal properties and photostability of the protein. Furthermore, it is possible to encapsulate different proteins in the different shells, in order to prepare multifunctional nano-carriers. Finally, the nanoparticles are applicable for intracellular delivery of their cargo. The incorporation of proteins into the particle matrix seems to increase delivery and reduce lysosomal degradation of the cargo. Therefore, the protein-doped silica nanoparticles constitute a promising novel tool for biomedical applications of nanoparticles, especially in the field of intracellular delivery of macromolecules.